青少年时期是抑郁等精神障碍的高发阶段,抑郁症已成为青少年致残的主要原因之一,但目前对青少年抑郁症的治疗效果有限,深入了解其神经生物学机制迫在眉睫。青春期应激与精神疾病相关,早期生活经历影响应激恢复力,母性关怀对后代行为有重要作用,但具体分子机制不明。2025年3月8日,武汉大学王志昊团队在Nature Communications杂志发表了题为“Maternal behavior promotes resilience to adolescent stress in mice through a microglia-neuron axis”的研究论文,该研究揭示了母性行为通过刺激小胶质细胞释放IGF-1,作用于神经元的IGF1R,激活mTORC1信号通路,抑制C/EBPβ的活性,从而减少Dnm1l的转录和DRP1的表达,防止线粒体功能障碍,增强青少年小鼠对压力的适应力。


图1 应激会致使青春期抑郁样小鼠mPFC的线粒体功能障碍
为明确线粒体功能障碍的哪些方面对青春期抑郁症的影响,研究人员向易感小鼠注射线粒体裂变抑制剂Mdivi1和过氧化物酶体增殖物激活受体-δ(PPAR-δ)激动剂GW0742。结果发现,Mdivi1能有效缓解AcSD诱导的社会回避和快感缺失行为,而GW0742无此效果(图2k-n)。已知Mdivi1是通过阻断DRP1(动力学相关蛋白1,基因名:Dnm1l)的自组装来阻止线粒体分裂的,进一步检测发现,Mdivi1可恢复AcSD诱导的线粒体功能障碍,改善线粒体形态,抑制青春期易感小鼠mPFC中DRP1表达,而GW0742则不能。
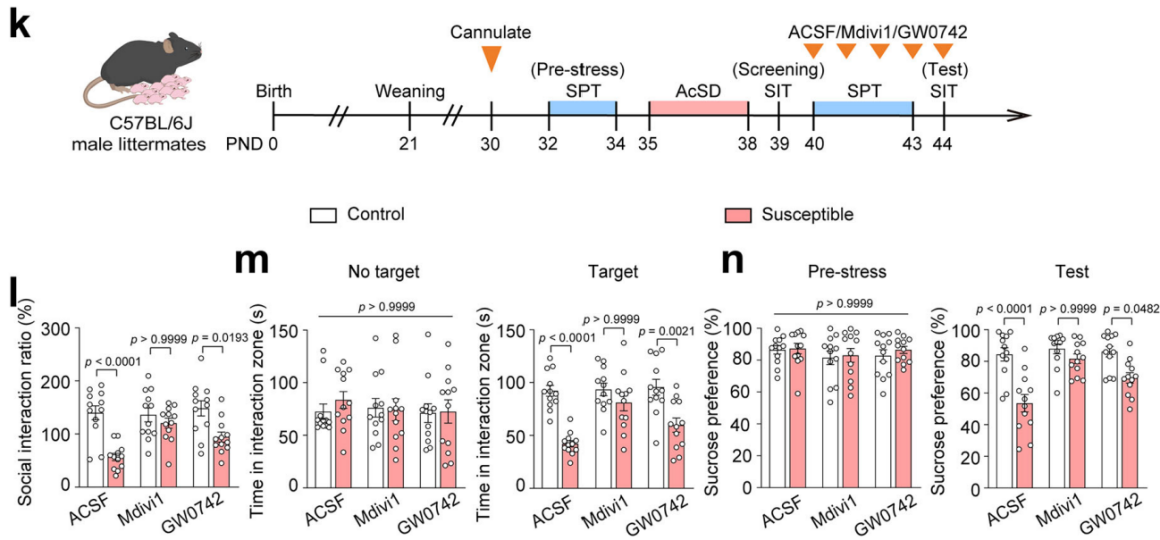
图2 Mdivi1可恢复AcSD诱导的线粒体功能障碍
研究人员进一步检测AcSD小鼠中DRP1的蛋白表达和mRNA水平,发现易感组小鼠mPFC中磷酸化和总DRP1蛋白表达以及Dnm1l mRNA水平均上调,且Dnm1l mRNA水平与社会互动比率呈负相关(图3a-c)。在不同时长的AcSD实验中,DRP1蛋白表达和mRNA水平随时间增加,4天AcSD会导致社会互动比率降低,经典抗抑郁药氟西汀可逆转这些变化。这些结果表明,DRP1与小鼠青春期抑郁样行为密切相关,在介导mPFC线粒体功能以及促进社会挫败应激后青春期小鼠抑郁样行为中发挥关键作用。人类样本的分析发现抑郁症患者背外侧前额叶皮质中Dnm1l的相对表达更高。此外,对AcSD小鼠脑切片进行了DRP1与NeuN、Iba1和GFAP标记物免疫共染,结果显示在青少年易感小鼠的mPFC中,DRP1与NeuN高度共定位(图3d-i)。重要的是,NeuN+细胞中的DRP1强度与社会互动比呈负相关(图2e),突出了mPFC神经元中DRP1在青少年小鼠抑郁样行为的作用。
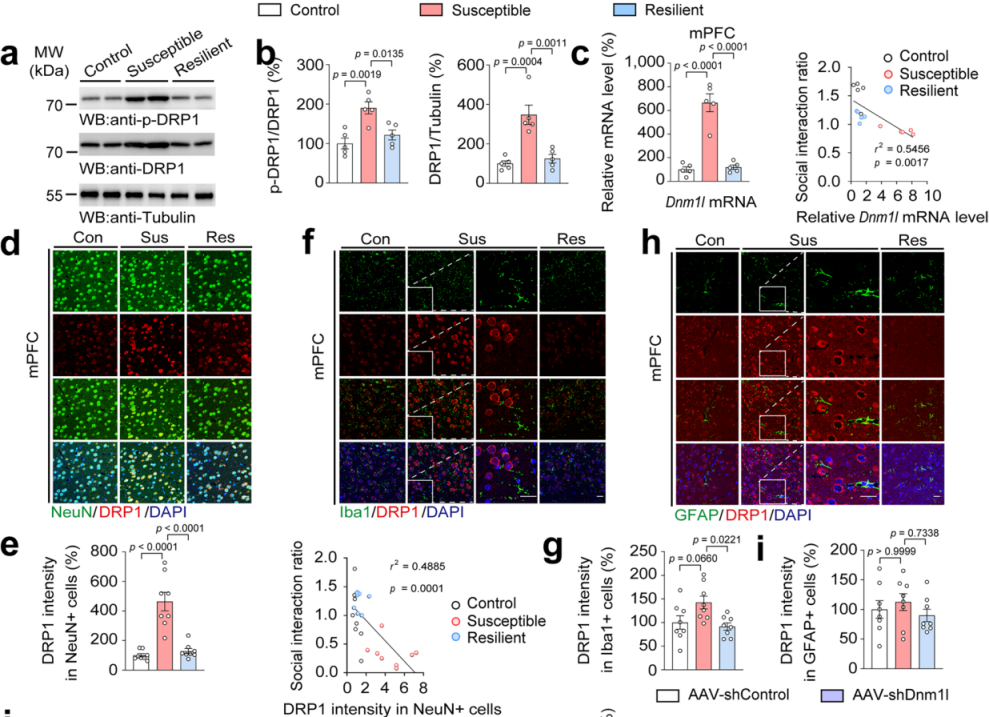
图3 易感小鼠mPFC神经元中DRP1蛋白水平和mRNA水平升高
在PND29(出生后第29天)时,将AAV-hSyn-shDnm1l-EGFP或AAV-hSyn-shControl-EGFP 注入野生型小鼠的mPFC以敲低神经元DRP1 (图4j),结果显示特异性降低DRP1不仅恢复了被AcSD破坏的线粒体功能,还显著提高了社会互动比率和蔗糖偏好(图4k-n)。这说明敲低DRP1能够改善因AcSD导致的线粒体功能障碍和抑郁样行为。应用 AAV-hSyn-Dnm1l-EGFP或 AAV-hSyn-EGFP在亚阈值社会挫败应激(SSD)小鼠mPFC神经元特异性过表达DRP1 (图4o),结果显示过表达DRP1诱导了SSD小鼠mPFC的线粒体功能障碍,过表达DRP1的小鼠社会互动和蔗糖偏好显著降低 (图4p-s)。这表明mPFC神经元中DRP1的增加会增强青春期小鼠的应激易感性。
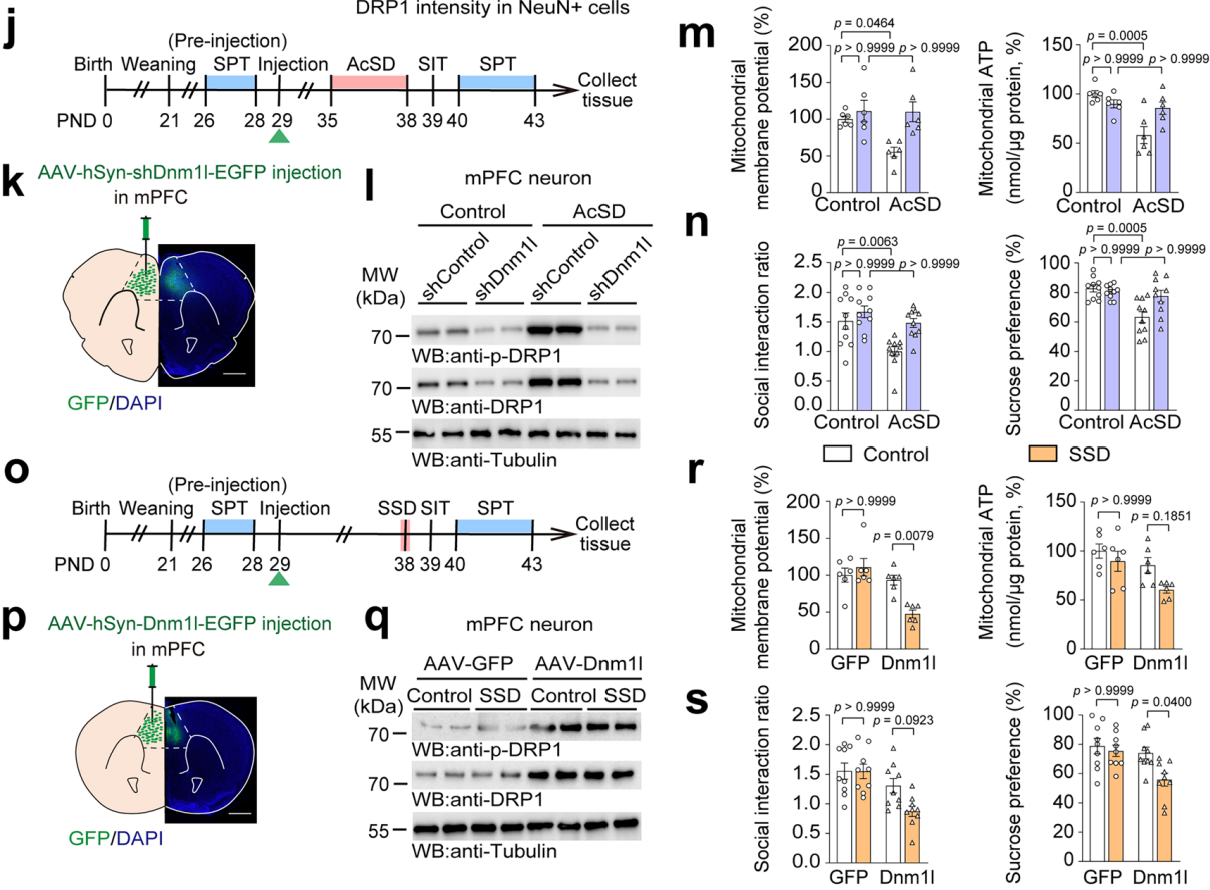
图4 mPFC神经元中DRP1参与调控青春期小鼠的应激易感性
基于ChIP-Atlas数据库从HeLa细胞中筛选出可能与Dnm1l启动子结合的转录因子,包括Gabpa、Ctcf、Rest和Cebpb(图5a),经检测在易感小鼠中只有Cebpb的mRNA水平显著高于对照组(图5b),且与社会互动比率呈负相关(图5c),蛋白分析也显示易感小鼠mPFC中磷酸化和总C/EBPβ水平增加(图5d)。荧光原位杂交(FISH)结合免疫染色分析表明,在易感小鼠的mPFC中,p-C/EBPβ和Dnm1l mRNA水平在细胞水平上均升高,并且p-C/EBPβ的强度与社会互动率之间存在显著的负相关(图5e-f),结果表明,p-C/EBPβ与Dnm1l mRNA高度相关(图5e-g)。免疫染色结果表明易感小鼠中C/EBPβ强度显著增加,并且与DRP1强度呈正相关(图5h-j)。这些发现表明,C/EBPβ可能是易感小鼠mPFC中DRP1的转录调节因子。
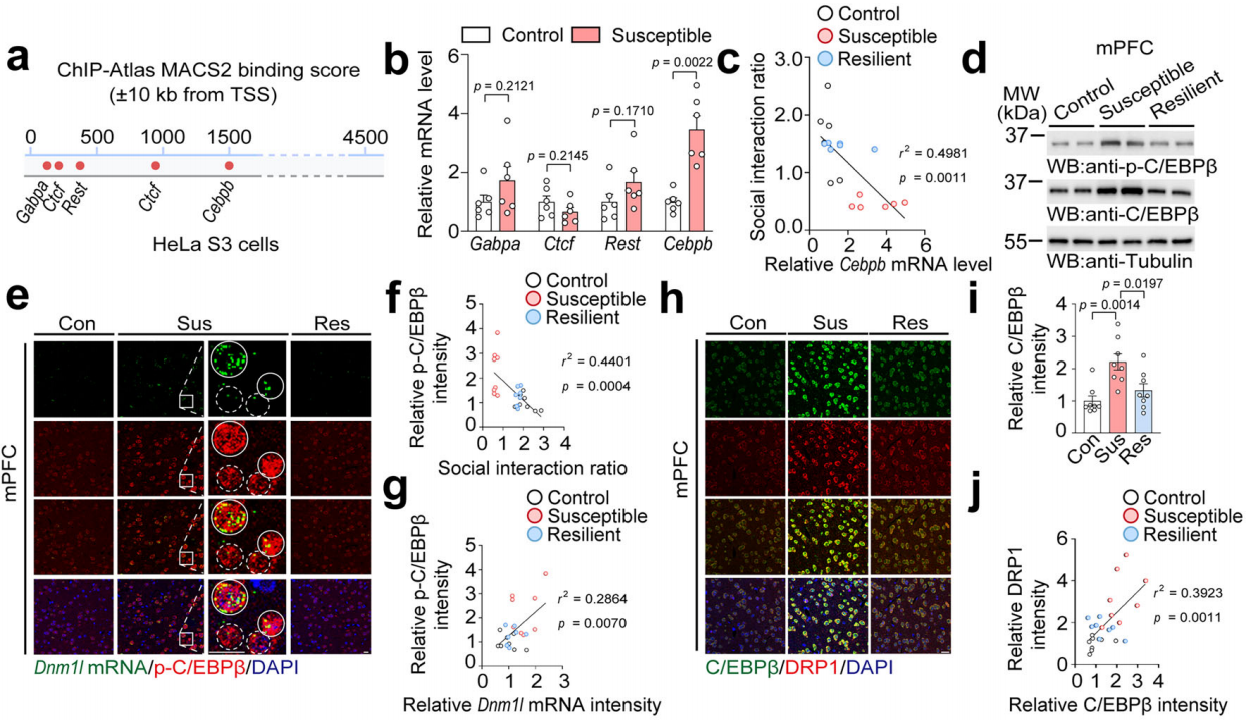
图5 易感小鼠中C/EBPβ强度显著增加并且与DRP1强度呈正相关
分析ChIP-seq数据发现C/EBPβ在Dnm1l启动子区域有高结合分数(图6k)。通过构建不同截断的Dnm1l启动子进行荧光素酶报告基因实验,确定了C/EBPβ在Dnm1l启动子上的主要结合位点(-1369至-1060片段)(图6l-o)。电泳迁移率变动分析(EMSA)和染色质免疫沉淀(ChIP)实验进一步证实C/EBPβ 能特异性结合Dnm1l启动子(图6p-q)。C/EBPβ有三种不同的蛋白亚型:即LAP*(肝脏激活蛋白)、LAP和LIP(肝脏抑制蛋白)。利用C/EBPβ LAP Tg小鼠(通过Thy1启动子过表达LAP亚型,以模拟神经元中过度激活的C/EBPβ)和原代小鼠皮层神经元实验,发现C/EBPβ敲除会降低DRP1的mRNA和蛋白水平,而在C/EBPβ LAP Tg 小鼠中则增加,表明C/EBPβ作为Dnm1l的转录激活因子能促进DRP1表达(图6r-w)。
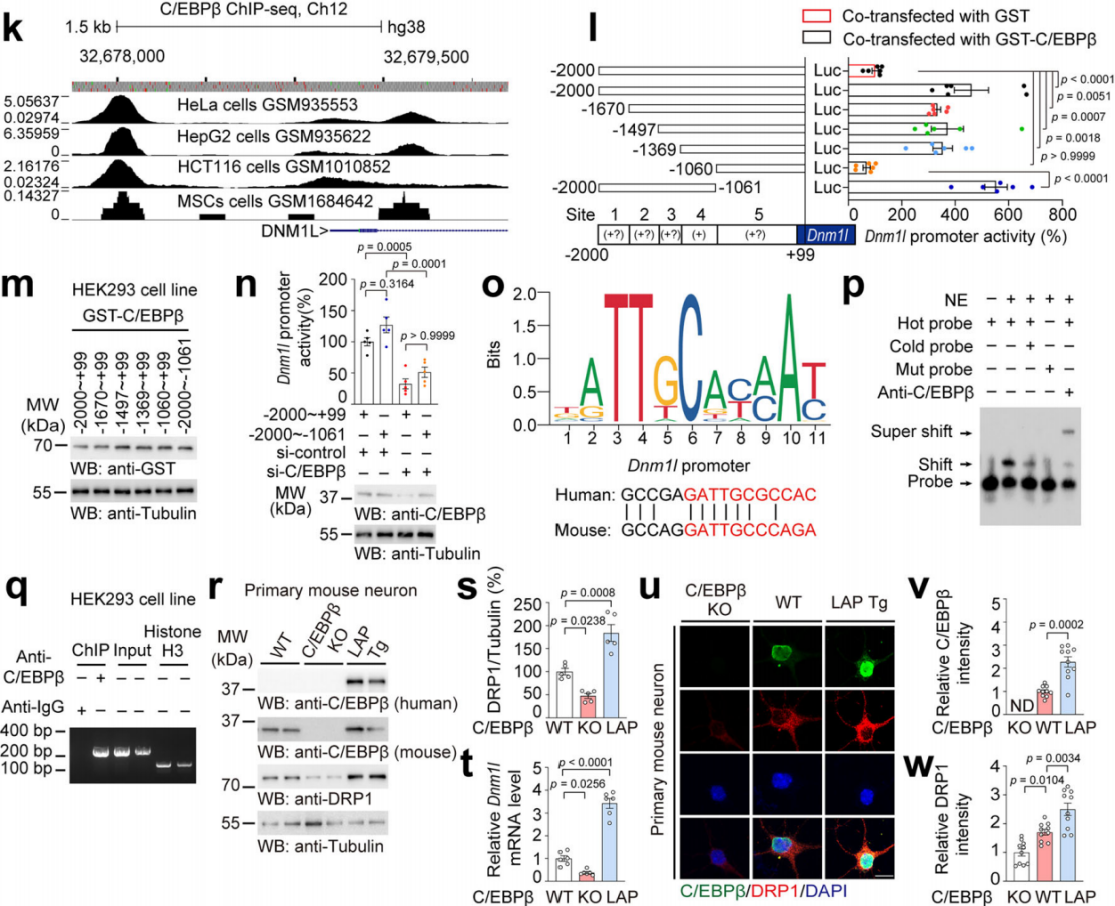
图6 C/EBPβ作为Dnm1l的转录激活因子能促进DRP1表达
通过免疫染色发现,在易感小鼠mPFC的NeuN+细胞中C/EBPβ强度最高(图7a-b)。此外,NeuN+细胞中的C/EBPβ强度与社会互动比值呈负相关(图7c)。这些结果表明,C/EBPβ主要在mPFC神经元中作为DRP1的转录因子。为了进一步阐明神经元C/EBPβ对青春期抑郁样行为的影响,繁殖了野生型(WT,C/EBPβ+/+)、C/EBPβ杂合敲除(C/EBPβ+/-)小鼠以及Thy1-人C/EBPβ LAP转基因(LAP Tg)小鼠。将AAV-hSyn-EGFP或AAV-hSyn-Dnm1l-EGFP注射到C/EBPβ+/-和WT小鼠的mPFC中,然后进行AcSD范式、行为评估和线粒体功能分析(图7d-g),结果显示C/EBPβ+/-小鼠mPFC神经元中的DRP1蛋白水平显著降低(图7f-g)。与WT小鼠相比,C/EBPβ+/-小鼠在经历AcSD后,线粒体膜电位和线粒体ATP水平改善,但过表达DRP1会逆转这些改善,表明在mPFC中敲除C/EBPβ可通过下调DRP1表达来减轻AcSD诱导的线粒体功能障碍(图7h-i)。此外,C/EBPβ+/-小鼠的社会互动比率和蔗糖偏好增加,而当过表达DRP1时这种增加消失,表明在mPFC中降低C/EBPβ可降低青春期小鼠的应激易感性(图7j-k)。
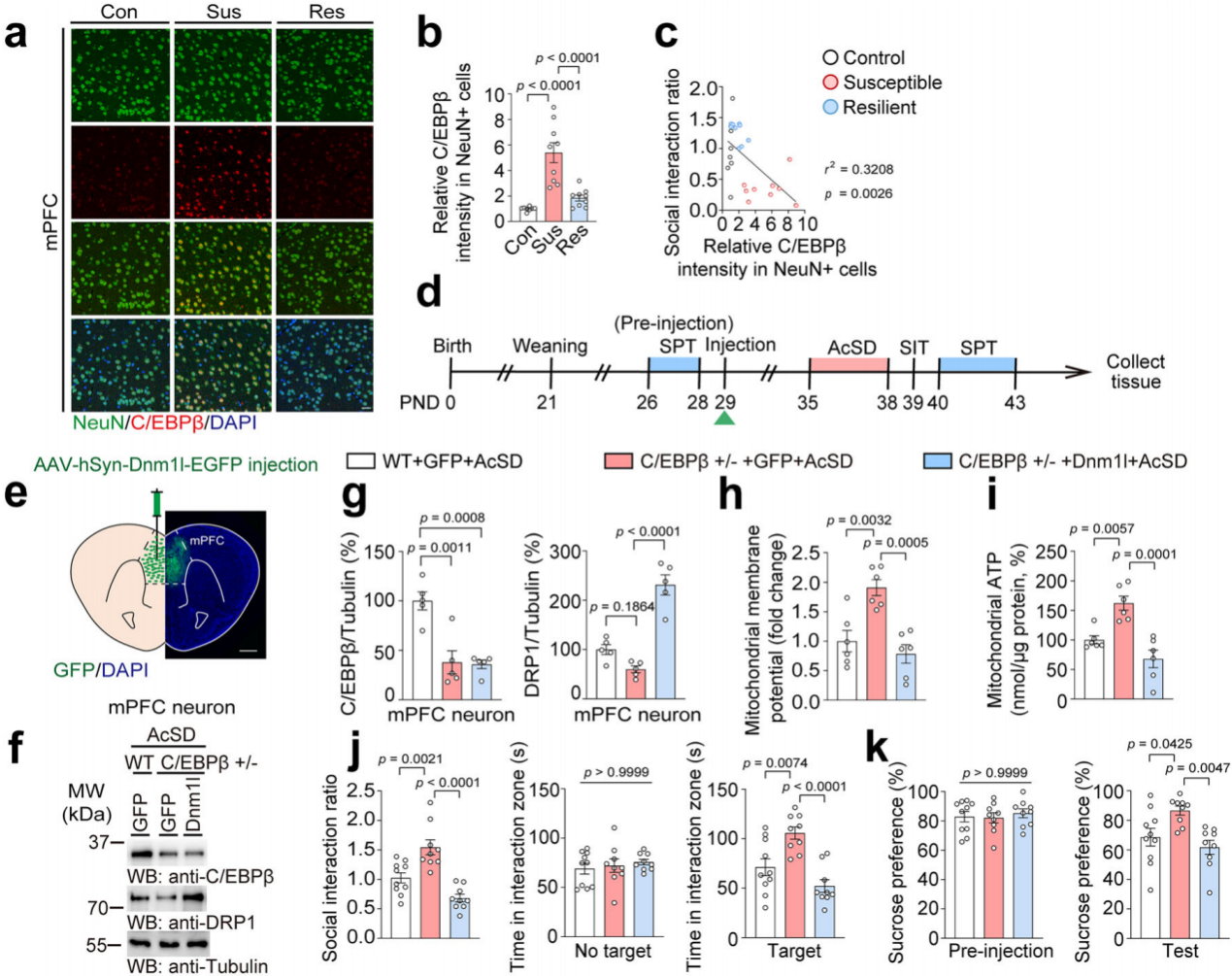
图7 下调mPFC中C/EBPβ可降低青春期小鼠的应激易感性
接下来,将AAV-hSyn-shDnm1l-EGFP或AAV-hSyn-shControl-EGFP注入LAP Tg和WT小鼠的mPFC中,然后进行亚阈值社会挫败应激(SSD)处理或对照处理(图8m),结果显示SSD处理后LAP Tg小鼠DRP1蛋白表达升高,但在WT小鼠中没有(图8l,n、o)。此外,敲低LAP Tg小鼠mPFC神经元中DRP1能减轻SSD诱导的的线粒体功能障碍,这意味着神经元C/EBPβ通过上调DRP1加剧了线粒体功能障碍(图8p-q)。同样,在仅接受SSD处理的LAP Tg小鼠中观察到社会行为和蔗糖偏好的缺陷,这表明过表达神经元C/EBPβ能增加了青春期小鼠出现抑郁样行为的易感性(图8r-s)。综上所述,这些数据表明C/EBPβ增强了mPFC神经元中DRP1的表达,并促成了青春期小鼠的抑郁样行为。
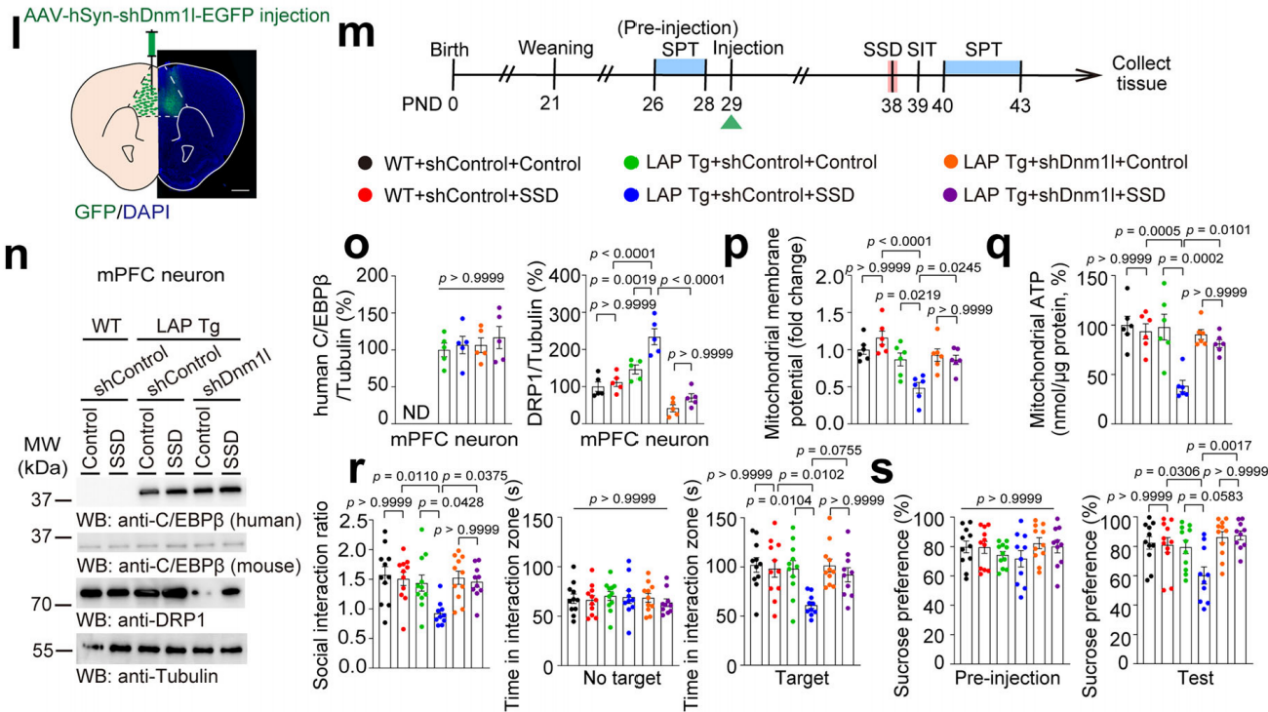
图8 C/EBPβ通过增强DRP1的表达促进青春期小鼠抑郁样行为
上述研究发现C/EBPβ过度激活会增加青少年小鼠对应激的易感性,但上游机制不明。流行病学和临床前研究表明早期生活应激源影响青少年抗压能力,“二次打击”模型提供了研究思路(图9a)。母婴分离(MS)作为经典模型可调节青少年应激易感性,因此从青少年慢性社会挫败模型转向母婴分离模型来研究促进神经元C/EBPβ活性的上游信号通路。母婴分离会改变青少年小鼠mPFC神经元中C/EBPβ活性和LAP/LIP比值(图9b-f),且在无出生后压力的易感小鼠中该比值也升高,提示该比值参与C/EBPβ激活及早期生活事件对青少年抑郁症的调节。此外,IGF-1可能通过IGF-1R/mTORC1信号通路调控C/EBPβ LAP/LIP比值(图9g),在经历母婴分离和无母婴分离经历的压力易感小鼠中均检测到低水平IGF-1,表明IGF-1在不同的时间线上将这两种压力源联系起来。
通过用IGF-1处理原代小鼠皮质神经元,发现IGF-1能增加IGF-1受体(IGF-1R)等相关蛋白磷酸化水平与总水平的比值(图9h-i),这表明IGF-1与其受体结合,激活mTORC1信号通路,降低LAP/LIP比值,下调了C/EBPβ活性,但未改变 C/EBPβ的总蛋白和mRNA水平,且与 Dnm1l mRNA 水平下降相关(图9i)。为了探究IGF-1在应激易感性中的作用,对青少年小鼠进行MS和AcSD实验,同时注射抗IGF-1或抗IgG阻断大脑IGF-1水平(图9j)。结果显示,抗IGF-1恢复了神经元LAP/LIP比以及DRP1的mRNA和蛋白水平,降低了处于压力下的青少年小鼠的社交互动比率等(图9k-m)。另外设置多组实验评估抗 IGF-1 抗体与AcSD单独和叠加作用,结果发现叠加作用加剧了这些变化。然而,10min×2 MS抑制了 C/EBPβ/DRP1 轴,并恢复了接受抗 IGF-1 抗体治疗或AcSD或两者兼有的青少年小鼠的行为表现。这表明,10min×2 MS诱导的 IGF-1 上调对于维持青少年小鼠的抗压能力很重要。
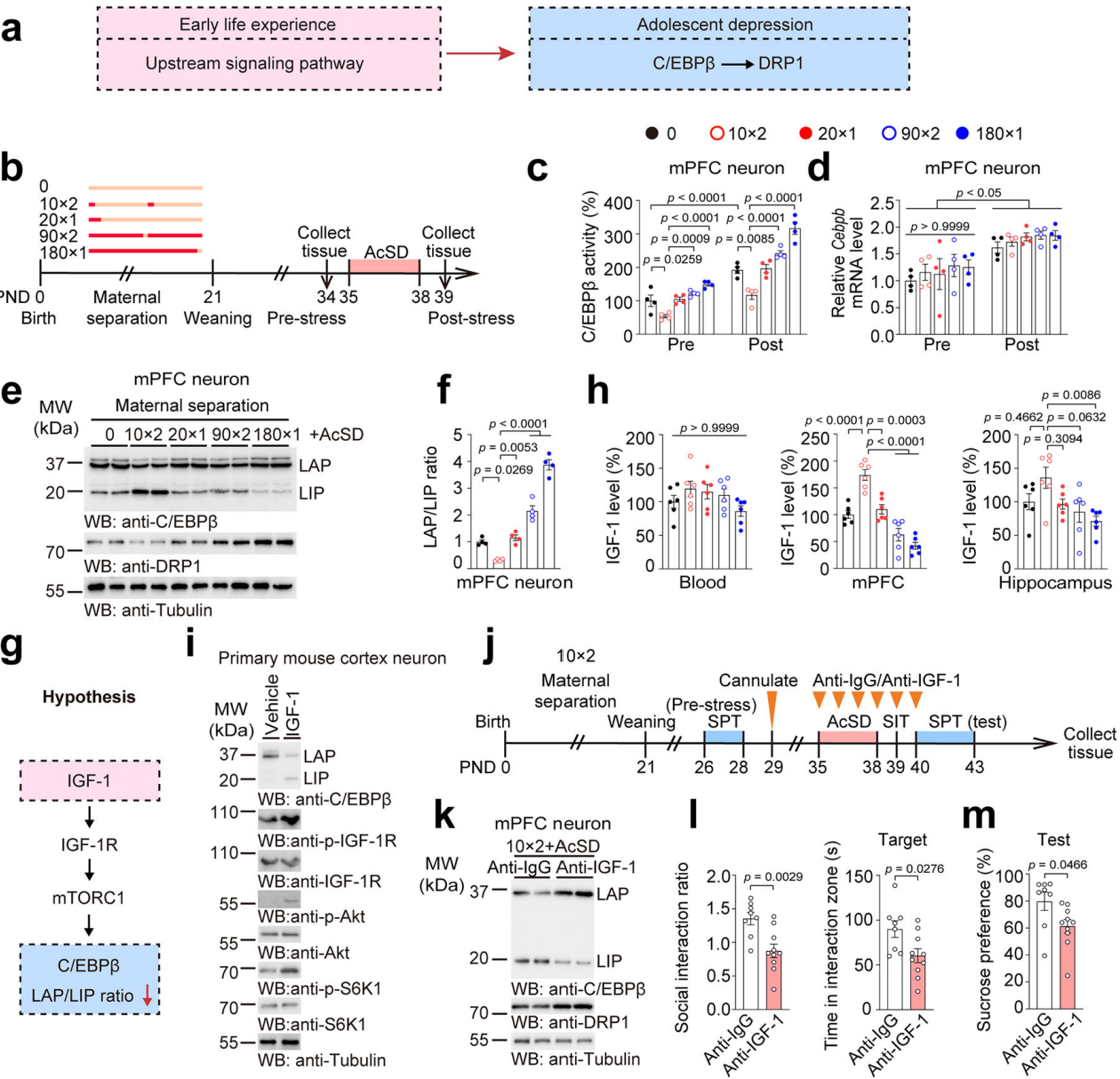
图9 10min×2 MS诱导的 IGF-1上调对维持小鼠抗压能力重要
筛选出易感小鼠后用IGF-1或对照药物处理,发现IGF-1可降低无母婴分离小鼠的LAP/LIP比值和DRP1蛋白表达(图10n-p),改善社交和蔗糖偏好缺陷(图10q-r);对经历长时间母婴分离的小鼠进行长时间IGF-1处理,可逆转相关指标变化,缓解社交回避和抑郁样行为。综上强调了大脑特异性IGF-1是介导小鼠应激易感性的关键分子介质,C/EBPβ-DRP1轴是重要下游通路。
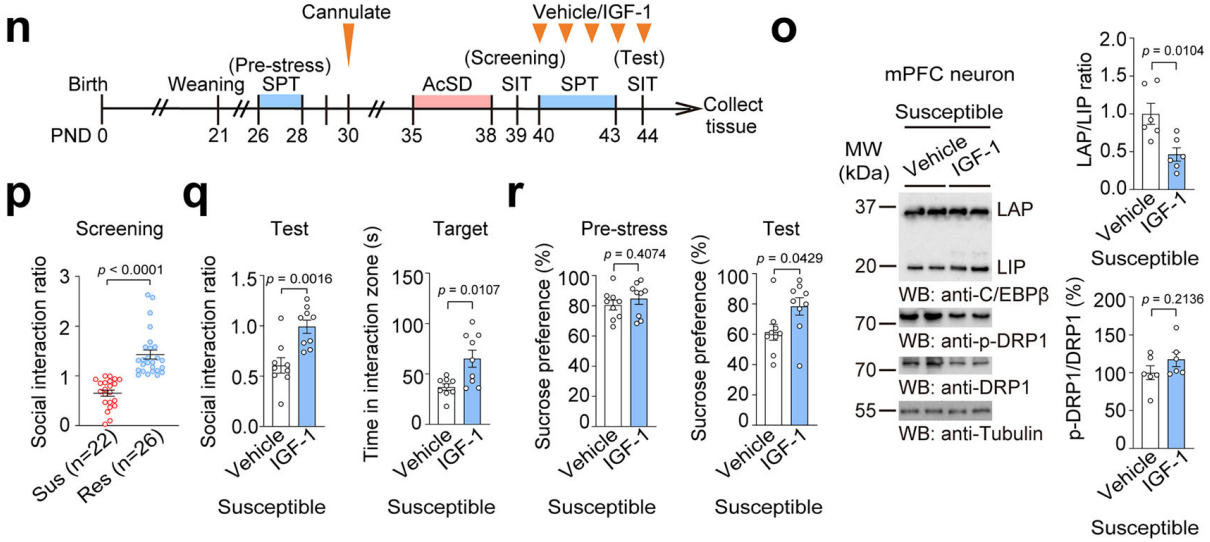
图10 用 IGF-1处理易感小鼠能改善相关指标和行为缺陷
母婴分离可能改变母性行为,研究人员量化母性行为后发现,10分钟×2母婴分离组母性行为样本点数增加,180分钟×1母婴分离组减少(图11a)。而且,母性行为样本点数与社会互动比率、mPFC的IGF-1水平呈正相关(图11b-d),说明母性行为在调节mPFC的IGF-1水平和保护小鼠免受应激易感性方面很重要。利用非哺乳母亲(LnL)模型增加对幼崽的母性行为,同时进行180分钟×1母婴分离和 AcSD范式实验(图11e)。结果显示,LnL组母性行为样本点数更高(图11f),能提升mPFC的IGF-1水平(图11g),降低LAP/LIP比值、DRP1蛋白表达和Dnm1l mRNA水平(图11h),改善长期母婴分离诱导的社会回避和蔗糖偏好缺陷(图11i-j),且不影响幼崽体重,表明增加的母性关怀可抑制C/EBPβ-DRP1轴。IGF-1在母性行为抗抑郁作用中的角色探究实验,进一步证明母性行为提高IGF-1水平通过调节LAP/LIP比值预防应激易感性。
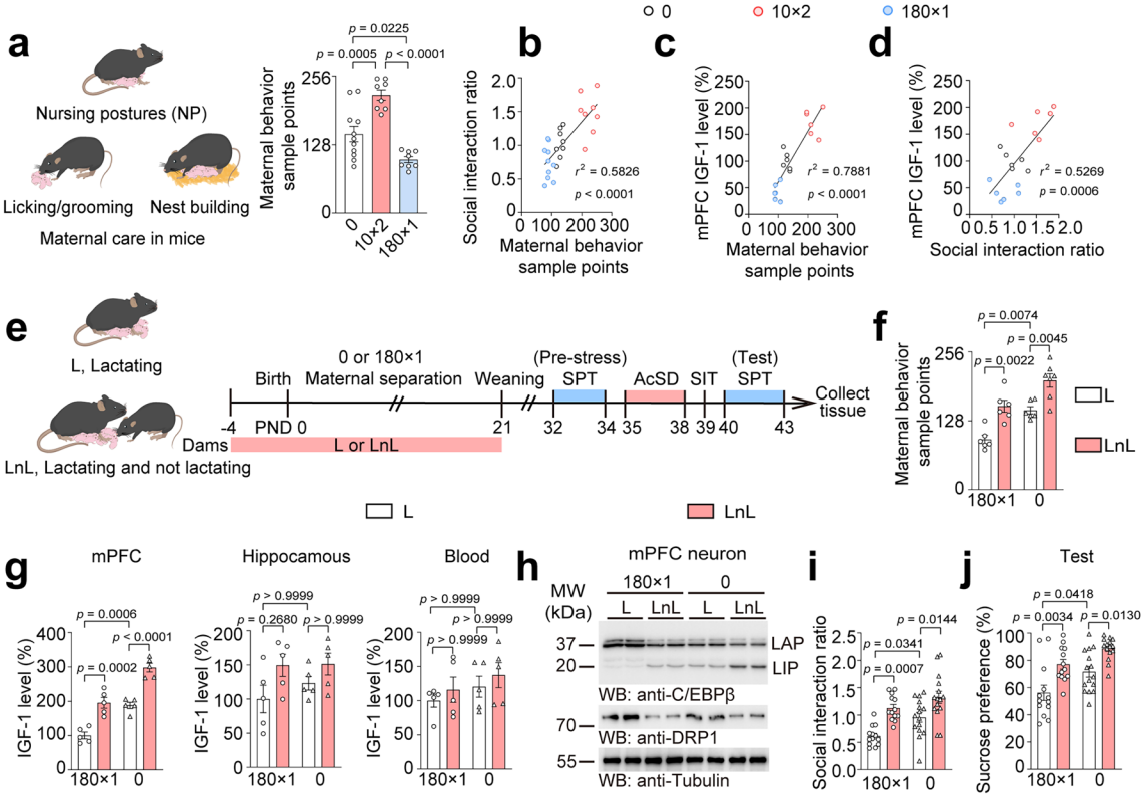
图11 母性行为提高IGF-1水平通过调节LAP/LIP比值预防应激易感性
据报道内侧视前区(MPOA)是与母性行为相关的重要脑区,尤其是MPOA中的Esr1⁺细胞在母性行为期间会被自然且优先激活。为了进一步阐释早期母性关怀与青春期应激易感性之间的关系,研究人员利用化学遗传学方法,在小鼠出生后0-8天抑制母鼠MPOA中Esr1⁺细胞活性,降低早期母性行为,之后进行10分钟×2的母婴分离或对照处理,再实施AcSD范式(图12k-l)。结果显示,CNO处理的母鼠的母性行为样本点数减少(图12m),幼崽mPFC中IGF-1水平降低(图6n),LAP/LIP比值和DRP1的mRNA及蛋白水平升高(图12o),青春期后代出现社会回避和蔗糖偏好缺陷,说明早期母性行为缺失会增加青春期应激易感性(图12p-q),且母性行为减少不影响幼崽养育质量。实验表明早期母性行为在调节青春期mPFC的IGF-1水平,增强小鼠应激恢复能力和预防抑郁样行为方面至关重要。(图12r)。
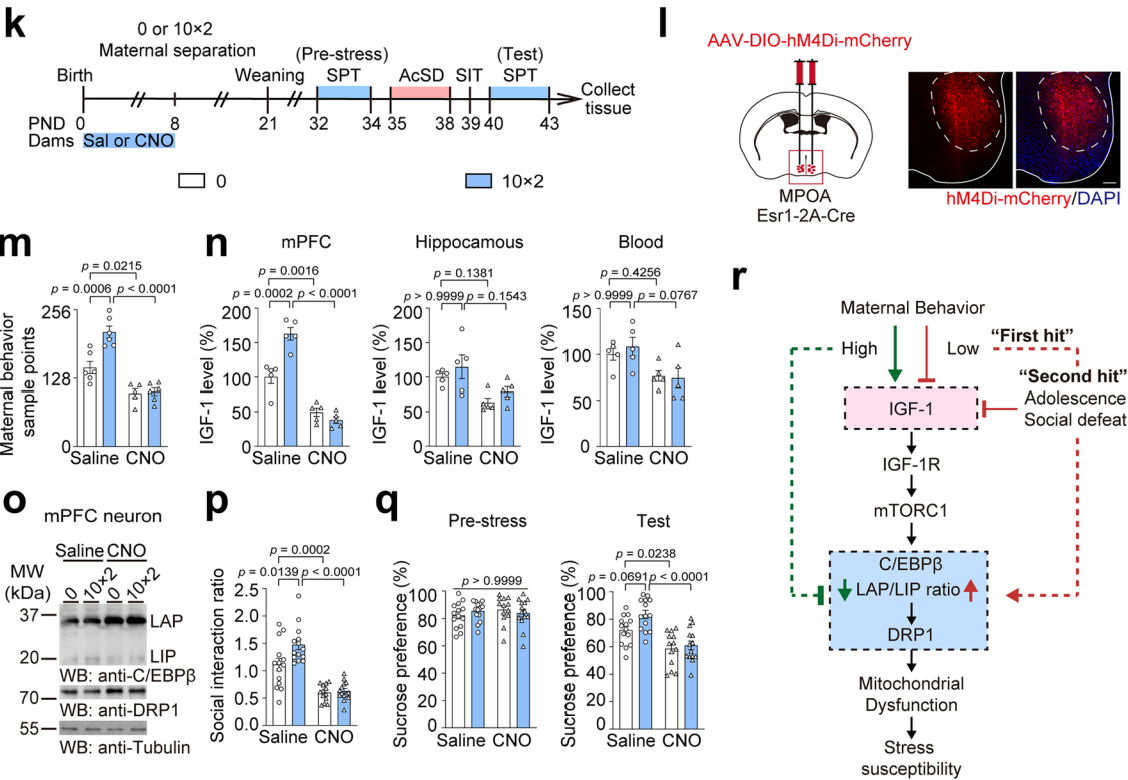
图12 早期母性行为缺失会增加青春期应激易感性
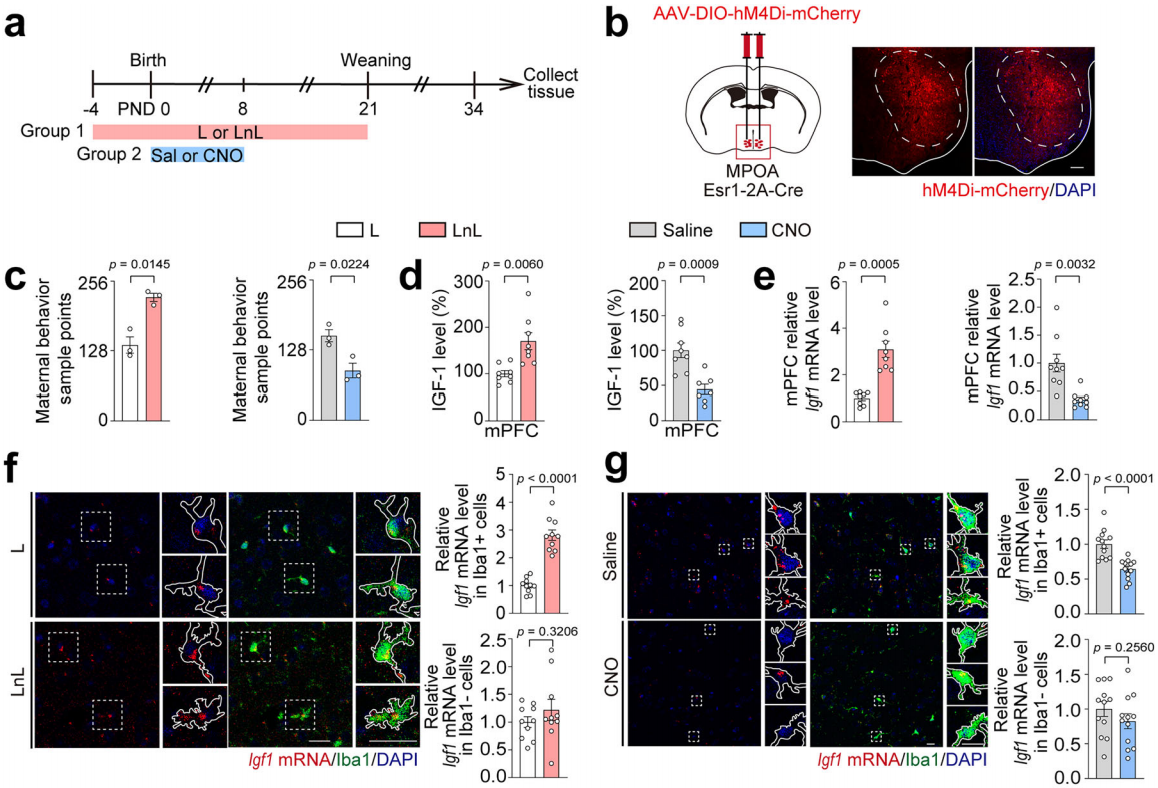
图13 小胶质细胞是幼崽响应母性行为产生IGF-1的主要来源
小胶质细胞是大脑中唯一表达集落刺激因子1受体(CSF1R)的细胞类型,给予CSF1R抑制剂PLX3397可有效清除小胶质细胞。为了研究小胶质细胞在母性行为介导的青春期小鼠 IGF-1 抗抑郁样效应中的作用,在对幼崽进行L或LnL处理以及AcSD范式后,给予幼崽 PLX3397(图14h)。结果显示,PLX3397处理阻断了LnL诱导的mPFC中IGF-1的升高和 C/EBPβ-DRP1的激活,从而导致DRP1相关的线粒体功能障碍(图14i-n)。这些结果表明,小胶质细胞缺失会降低IGF-1水平,进而激活C/EBPβ 对 Dnm1l的转录活性,导致线粒体功能障碍。行为测试表明,PLX3397 处理消除了增加的母性行为对AcSD的保护作用,导致社会互动减少、在互动区域停留的时间缩短以及蔗糖偏好受损(图14o-p)。结合小胶质细胞表型实验,表明mPFC中的抗炎性小胶质细胞在母性关怀的刺激下产生并分泌IGF-1,通过改善青春期小鼠神经元的线粒体功能来预防抑郁样行为。

图14 抗炎性小胶质细胞在青春期抑郁样行为中的保护作用
为深入了解小胶质细胞与神经元之间的相互作用,研究人员利用Cx3cr1-Cre小鼠和AAV-MG1.2-DIO病毒来调控小胶质细胞来源的IGF-1水平 ,并通过AAV病毒在神经元中特异性过表达C/EBPβ异构体。选择性抑制经LnL处理的幼鼠mPFC神经元中由小胶质细胞产生的IGF-1水平,并过表达LIP异构体(图15a)。实验结果显示,敲减小胶质细胞的 IGF-1 增加了 LAP/LIP 比值和 DRP1 蛋白表达(图15b-e),促进了线粒体功能障碍(图15f-g),导致社会回避和蔗糖偏好缺陷;而神经元中 LIP 异构体的过表达则逆转了这些变化,减轻了抑郁样行为(图15h-i)。
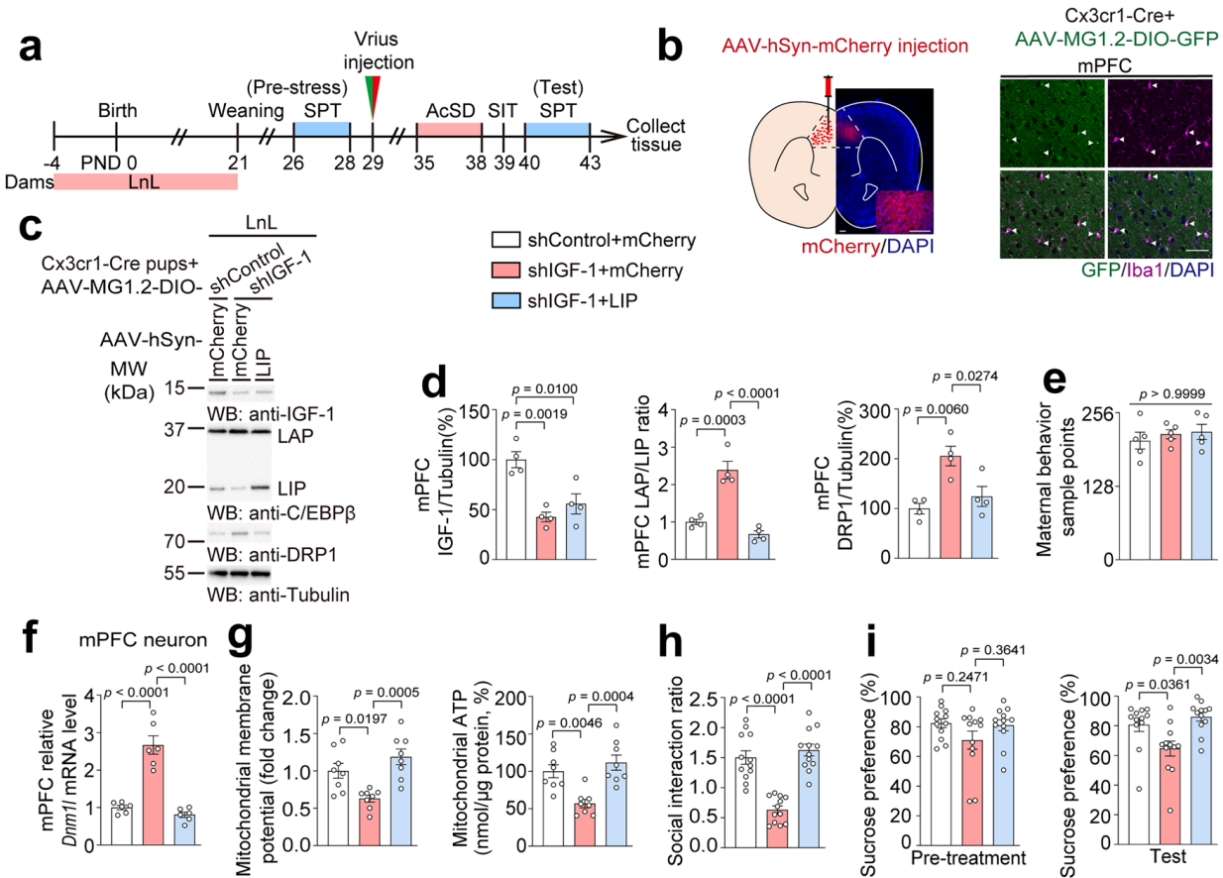
图15 抑制小胶质细胞来源的IGF-1并过表达LIP异构体
另外,在 CNO 处理的母鼠所产幼崽的小胶质细胞中补充 IGF-1,同时在 mPFC 神经元中过表达 LAP 异构体(图16j)。结果表明,补充 IGF-1 抑制了 LAP/LIP 比值,降低了 DRP1 蛋白表达(图16k-n),上调了线粒体 MMP 和 ATP 水平(图16o-q),改善了社会回避和蔗糖偏好缺陷(图16r);但神经元中过表达 LAP逆转了这些效果。综上所述,这些结果表明小胶质细胞与神经元之间的相互作用通过 IGF-1-C/EBPβ-DRP1 信号通路,在介导应激易感性方面发挥着重要作用。
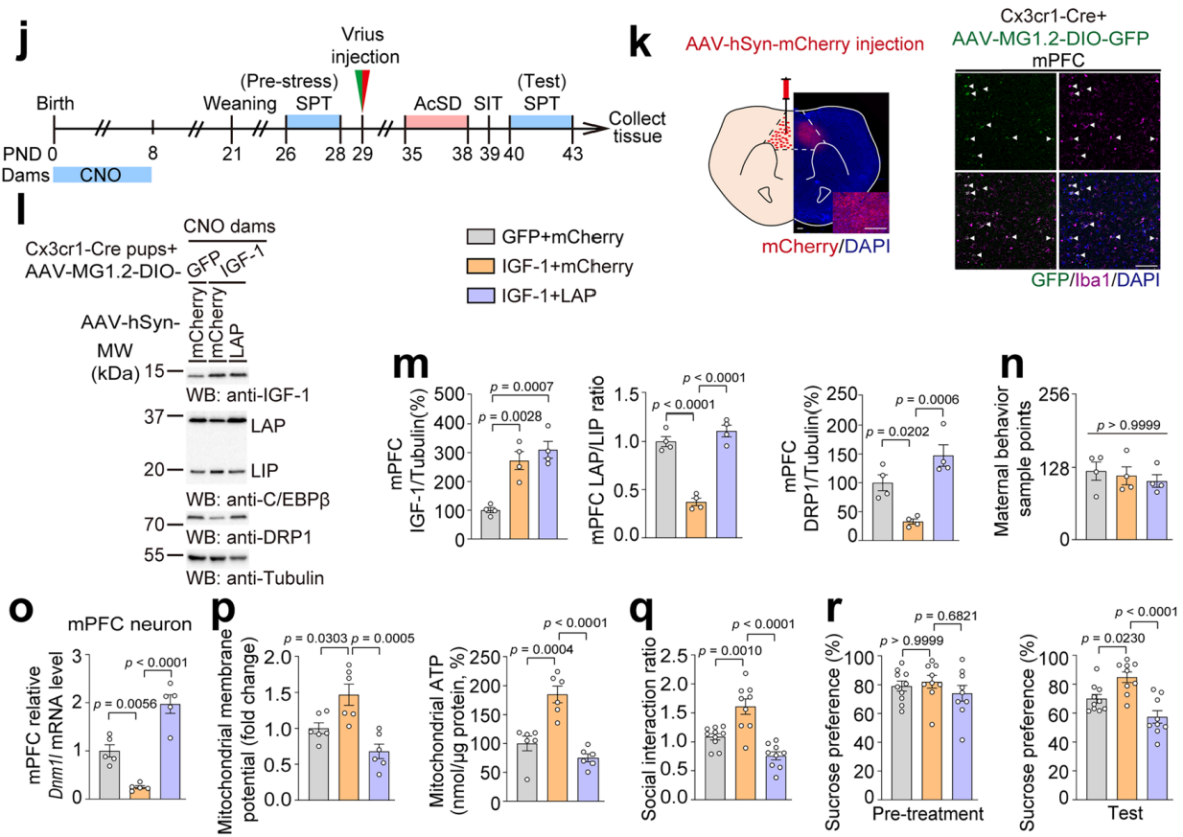
图16 补充小胶质细胞来源的IGF-1并过表达LAP异构体
本研究发现母性行为通过小胶质细胞-神经元轴,即IGF-1-C/EBPβ-DRP1信号通路,调节线粒体功能,影响青春期小鼠应激易感性。明确了社会应激会激活青春期小鼠mPFC神经元中C/EBPβ,进而上调Dnm1l,导致线粒体功能障碍,增加应激易感性;而母性行为刺激产生的IGF-1可抑制该通路,起到保护作用。这些结论深化了对青春期应激相关神经生物学机制的理解。文章还证实了早期母性行为对预防青春期抑郁样行为至关重要,为青少年抑郁症的早期干预提供了科学依据。提示可通过改善早期生活环境、加强母性关怀等方式,降低青少年抑郁症的发生风险,对公共卫生领域制定相关干预策略有重要指导意义。
本文使用的工具病毒布林凯斯生物科技有限公司均可提供:
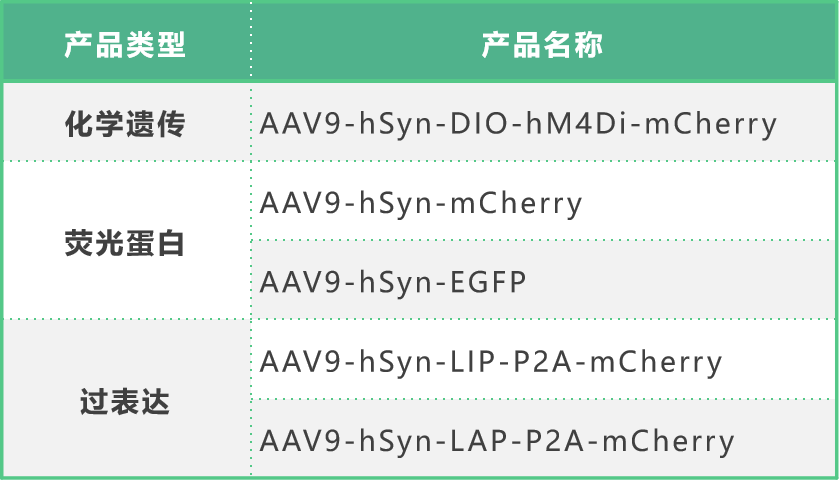
同时布林凯斯也可提供各类定制服务请联系小布:18971216876(微信同号)或者咨询所在区域的销售经理获取更多信息。




地址:-
地址:-
