稀疏标记 (sparse labeling)
简介
不同神经元个体间形态结构千差万别,轴树突纤维彼此缠绕,因此常规标记很难揭示群体中单个神经元清晰完整的形态结构以及彼此间的组织关系,这制约了人们理解神经元精细形态结构和神经环路组织构架。稀疏标记方法的开发有助于对大脑神经元进行数目稀少且高亮的标记,以研究其中单个神经元的局部树突形态和全脑完整轴突投射。
稀疏标记通过重建单个神经元全细胞形态的投射模式,精确解析单个神经元在神经环路中的形态结构。因此,该技术不仅能够重构全脑范围的单神经元投射形态,还能解析单神经元信息输入输出位点,为了解单神经元如何形成复杂多变的神经环路提供支持。
稀疏标记的方法
常用稀疏标记方法主要包括病毒标记以及转基因小鼠遗传学标记等,其中病毒标记方法以重组腺相关病毒(rAAV)的应用最为广泛。
布林凯斯采用
双包装质粒鸡尾酒法制作rAAV稀疏标记载体。该方法将
双核心表达质粒预混合后,与携带Rep基因(AAV复制蛋白)、Cap基因(AAV血清型结构蛋白)的包装质粒,携带腺病毒辅助包装基因的包装质粒混合后共同转染HEK293细胞,进一步纯化获得单一rAAV集合。
其他常用的稀疏标记方法有:
1. Cre剪切酶依赖:将高倍稀释表达Cre剪切酶的rAAV病毒与Cre剪切酶依赖表达荧光蛋白的rAAV病毒混合后注射小鼠大脑特定脑区。
2. Cre-ER转基因小鼠他莫昔芬依赖:通过控制他莫昔芬药物剂量,在Cre-ER转基因小鼠中,诱导特定细胞类型神经元的稀疏标记。
3. 四环素诱导系统泄露机制依赖:该方法利用四环素诱导表达系统的泄露机制,失效特定或非特定细胞类型神经元。
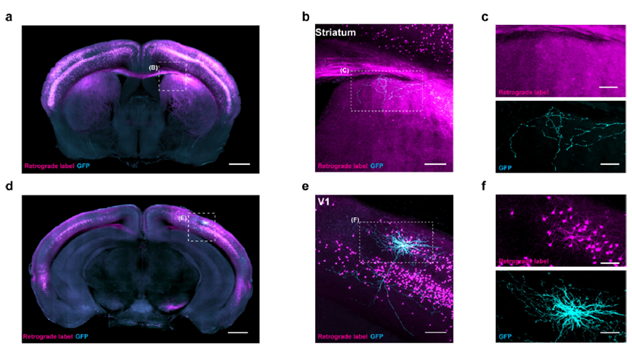
4. 突触末梢浸入限定感染数目:该方法通过逆向运输表达Cre剪切酶的病毒,控制神经元轴突末梢侵入限定感染数目,启动上游神经元中Cre剪切酶依赖的荧光蛋白表达,实现游目标脑区神经元的稀疏标记。
图1. 逆向稀疏标记示例(Han Y et al. Nature, 2018)
Brain Case稀疏标记的优势
①稳定性高:减少了不同病毒间的相互排斥,提高了共感染效果,使得每次病毒注射标记的神经元数量相对稳定。
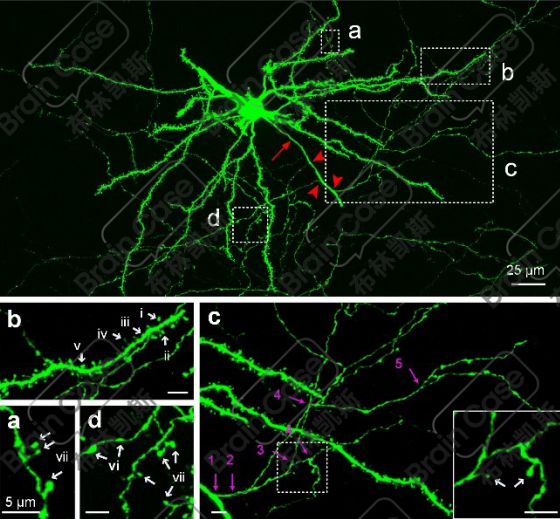
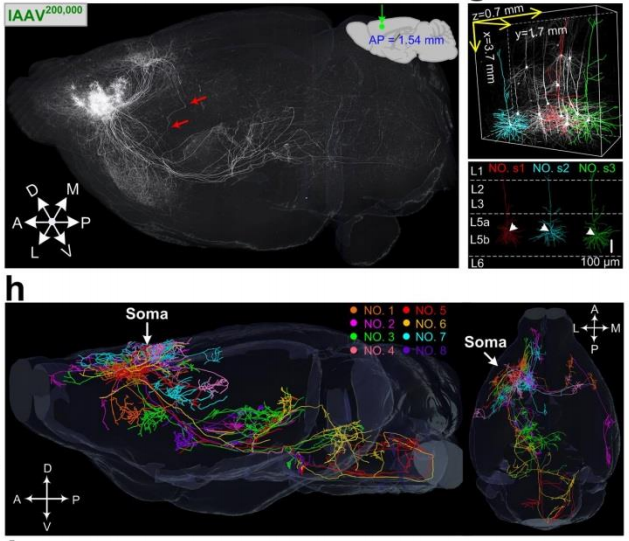
②高亮度标记:高亮度标记单个神经元精细形态,不仅仅是胞体,长程的轴突也可以被高亮地标记出来。
③稀疏度可控:通过调整Cre或Flp重组酶的比例,控制被标记神经元的数目,同时保持标记神经元的亮度不变。
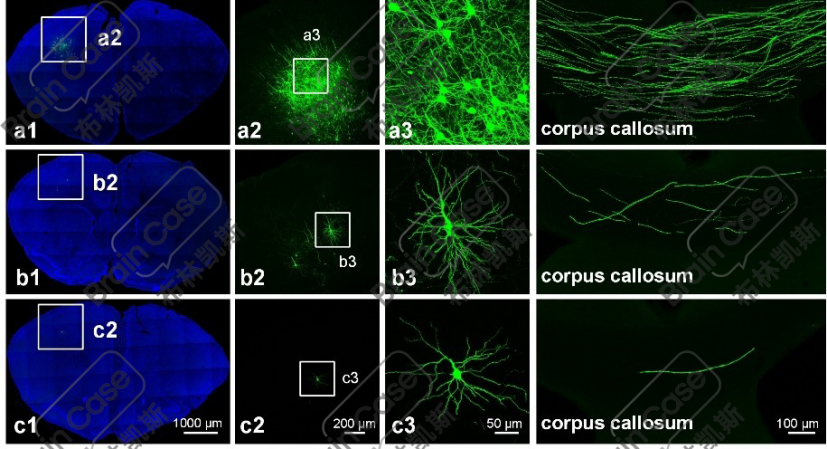
图2. 不同比例质粒混合进行注射的成像结果示意图
④细胞类型特异性:结合特定类型的Cre转基因动物或者特定类型神经元的启动子,可以高效标记特定类型的神经元。
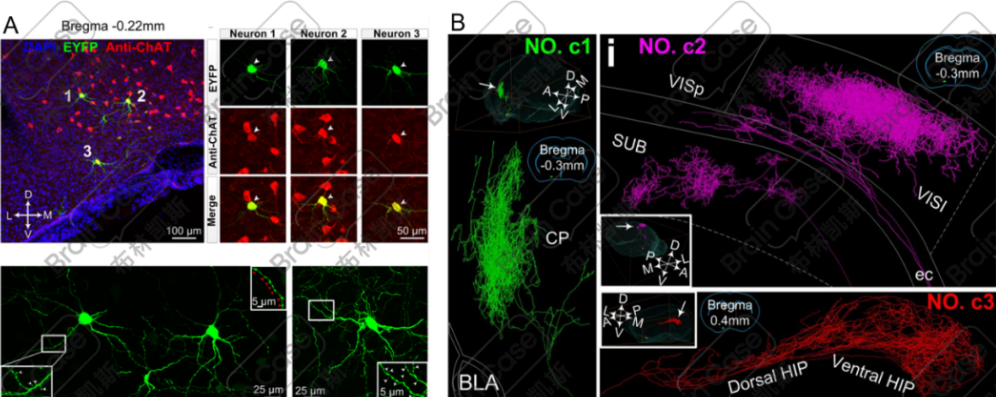
⑤可与多项技术联用,例如荧光显微光学切片断层成像(fMOST)、组织透明化、激光共聚焦等,以用于全脑成像和单细胞重建。
Brain Case稀疏标记产品
如果您的实验需要:神经元形态和结构成像;描绘树突精细结构;重建单神经元的全脑投射;解析单神经元输入输出位点即可考虑使用稀疏标记系列产品。并且,可以根据实验需求选择合适的产品。
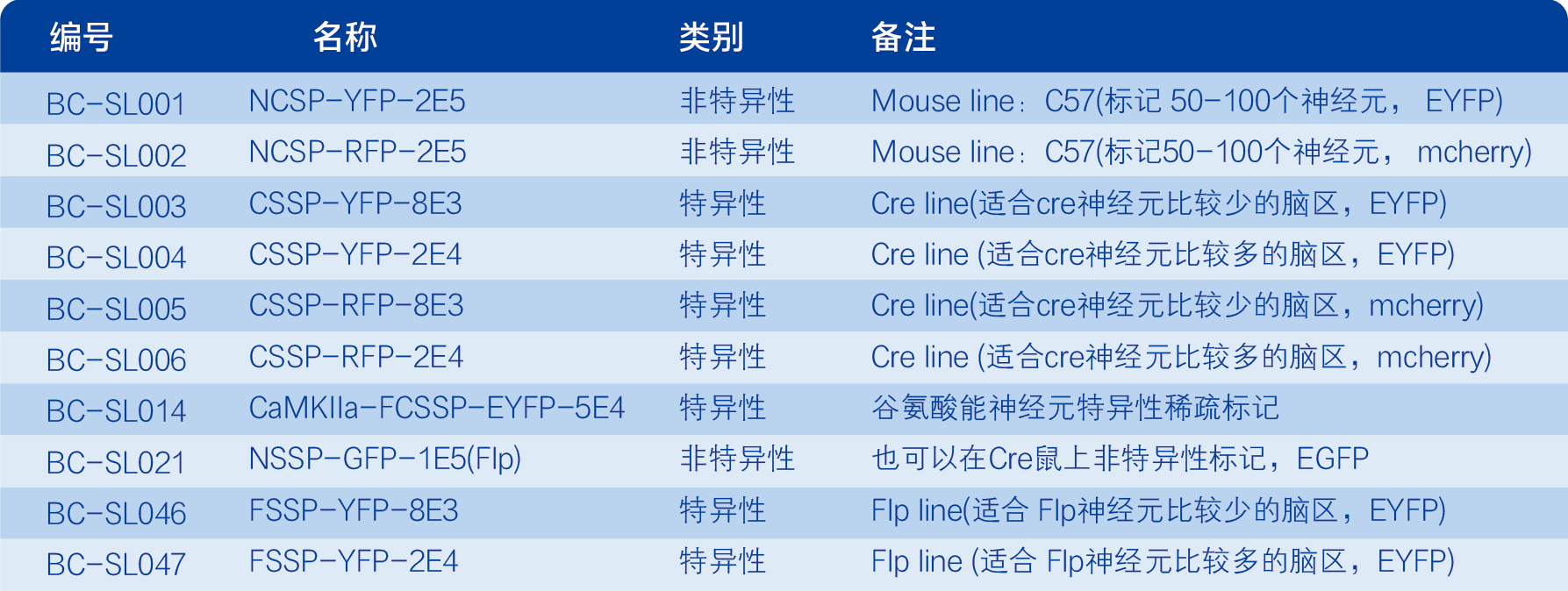
①根据实验动物,选择合适特异性类别。如果实验动物为 Cre小鼠,即可选择特异性稀疏标记产品,如BC-SL003或BC-SL004等;WT小鼠,可选择非特异性稀疏标记产品,如BC-SL001或BC-SL002。
②根据实验需求,选择合适的荧光蛋白。如果需要绿色标记,则选择YFP(EYFP)产品;需要红色标记,则选择RFP(mCherry)产品。
③根据标记神经元数量,选择合适的配比。如果使用Cre小鼠,请根据Cre神经元的密集程度选择合适的产品:Cre神经元比较稀疏的脑区,可选择BC-SL003或BC-SL005;Cre神经元比较密集的脑区,可选择BC-SL004或BC-SL006。
同时,该系列产品可根据您的实验需求进行特别定制。如果您对稀疏标记系列产品感兴趣,可以联系小布18971216876(微信同号)了解详情或下单订购。
案例展示
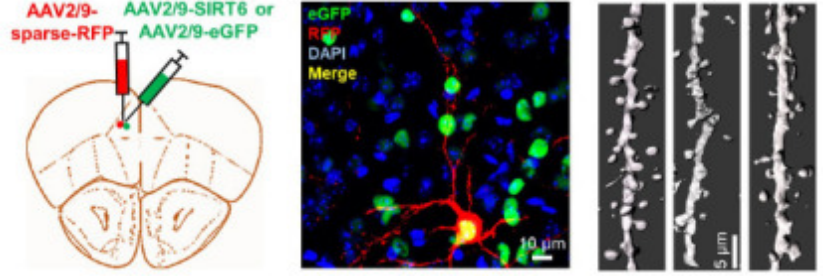
实验动物:C57BL/6J小鼠
病毒载体:BC-SL002
实验方法和结果:共包装 AAV-CMV-cre 质粒和 AAV-DIO-mCherry 质粒制作成单一rAAV集合,注射C57BL/6小鼠的单侧PrL中,稀疏标记神经元。
Zhang Z, Wang J, et al, Neural Regen Res, 2023
(PMID: 37282476)
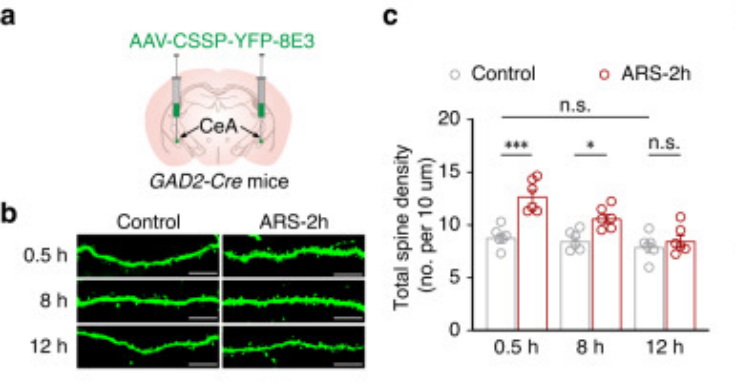
实验动物:GAD2-Cre小鼠
病毒载体:BC-SL003
实验方法和结果:共包装 AAV-CMV-DIO-Flp 质粒和 AAV-FDIO-EYFP 质粒制作成单一rAAV集合,注射GAD2-Cre小鼠双侧 CeA 中,特异性稀疏标记 GABACeA 神经元。
Cao P, Jin Y, et al, Nat Commun, 2024
(PMID: 38200023)
客户发表文章
【1】Wang L, Yang Z, Satoshi F, Prasanna X, Yan Z, Vihinen H, Chen Y, Zhao Y, He X, Bu Q, Li H, Zhao Y, Jiang L, Qin F, Dai Y, Zhang N, Qin M, Kuang W, Zhao Y, Jokitalo E, Vattulainen I, Kajander T, Zhao H, Cen X. Membrane remodeling by FAM92A1 during brain development regulates neuronal morphology, synaptic function, and cognition. Nat Commun. 2024 Jul 23;15(1):6209. doi: 10.1038/s41467-024-50565-w. PMID: 39043703; PMCID: PMC11266426.
【2】Chen D, Lou Q, Song XJ, Kang F, Liu A, Zheng C, Li Y, Wang D, Qun S, Zhang Z, Cao P, Jin Y. Microglia govern the extinction of acute stress-induced anxiety-like behaviors in male mice. Nat Commun. 2024 Jan 10;15(1):449. doi: 10.1038/s41467-024-44704-6. PMID: 38200023; PMCID: PMC10781988.
【3】Xie G, Gao X, Guo Q, Liang H, Yao L, Li W, Ma B, Wu N, Han X, Li J. Cannabidiol ameliorates PTSD-like symptoms by inhibiting neuroinflammation through its action on CB2 receptors in the brain of male mice. Brain Behav Immun. 2024 Jul;119:945-964. doi: 10.1016/j.bbi.2024.05.016. Epub 2024 May 15. PMID: 38759736.
【4】Liu M, Gao Y, Xin F, Hu Y, Wang T, Xie F, Shao C, Li T, Wang N, Yuan K. Parvalbumin and Somatostatin: Biomarkers for Two Parallel Tectothalamic Pathways in the Auditory Midbrain. J Neurosci. 2024 Mar 6;44(10):e1655232024. doi: 10.1523/JNEUROSCI.1655-23.2024. PMID: 38326037; PMCID: PMC10919325.
【5】Cheng J, Ma X, Li C, Ullah R, Wang X, Long J, Yuan Z, Liu S, Fu J, Chen Z, Shen Y, Zhou YD. Diet-induced inflammation in the anterior paraventricular thalamus induces compulsive sucrose-seeking. Nat Neurosci. 2022 Aug;25(8):1009-1013. doi: 10.1038/s41593-022-01129-y. Epub 2022 Aug 1. PMID: 35915173.