狂犬病毒(Rabies Virus,RV)属于弹状病毒科(Rhabdoviridae)狂犬病毒属(Lyssavirus),其基因组是长约12 kb的单股负链RNA,由3′端至5′端依次排列着N、P、M、G和L共5个狂犬病毒的结构基因,分别编码核蛋白、磷蛋白、基质蛋白、糖蛋白和转录酶大蛋白(如图1 )。
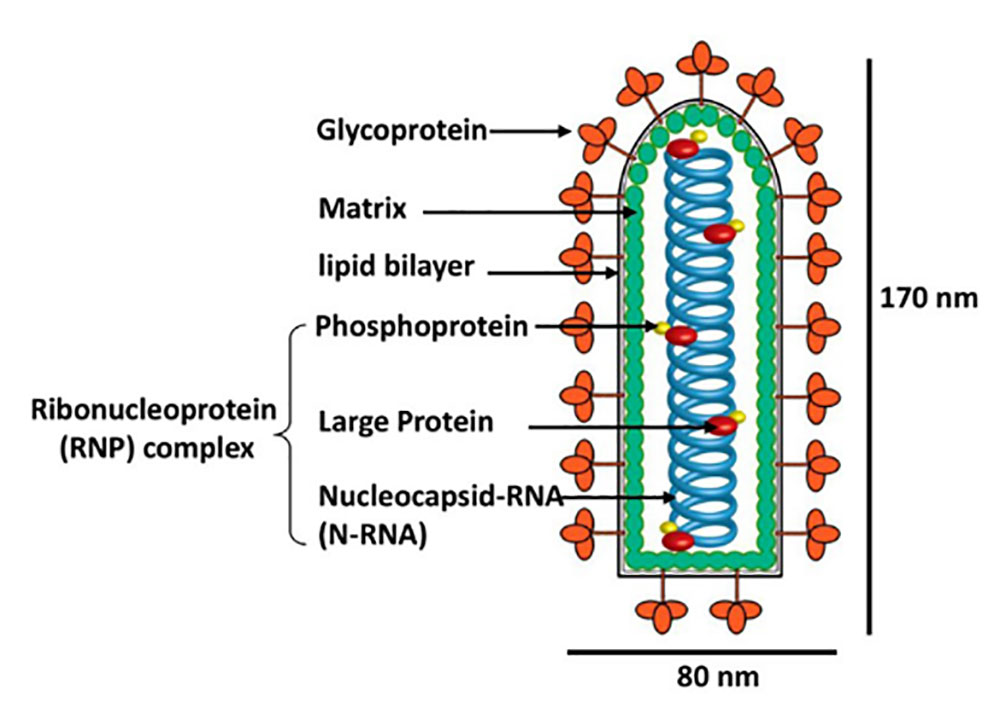
图1. RV粒子示意图(图片来自whelanlab网站)
RV在宿主体内,病毒糖蛋白G介导病毒粒子与细胞表面受体结合,通过内吞作用进入细胞。随后通过pH介导膜融合,促使RV基因组释放到细胞质中。感染病毒粒子内包装的L、N、P共同构成的聚合酶复合物以基因组负链RNA为模板启动转录表达,复制,组装,出芽产生子代病毒。(如图2)
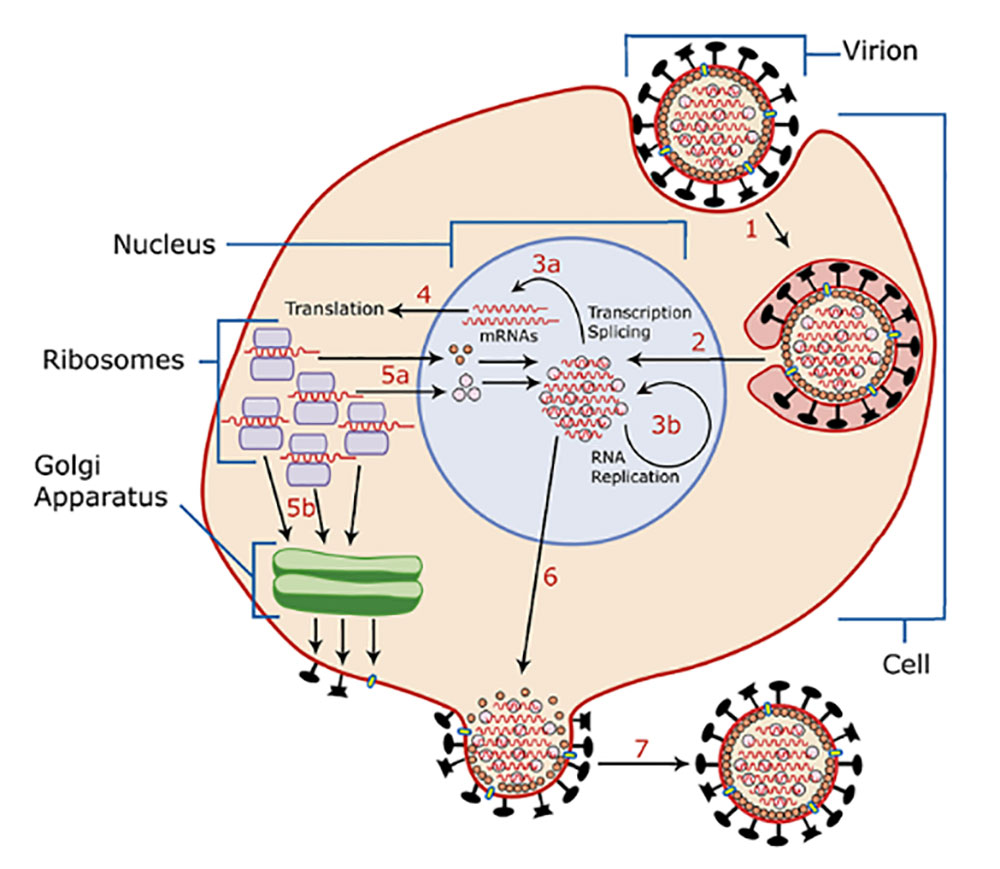
图2. 狂犬病毒生命周期示意图(图片来自维基百科)
野生型RV是人畜共患的狂犬病的致病因子,在神经系统中具有传播属性,对人和动物均具有较高致病性。因此本公司RV产品以狂犬病毒疫苗株(SAD B19或CVS-N2c)且缺失G蛋白的复制缺陷型RV (RV-SAD-ΔG/ RV-CVS-ΔG) 为骨架改造而来。随着反向遗传学手段的成熟,经过改造的RV,可用于神经回路研究。RV感染中枢系统后,主要标记神经元,几乎不标记胶质细胞;被感染的神经元在一定时间内(7-12天)几乎不发生明显病变及裂解。目前RV系统可分为逆向标记系统和逆向跨单突触系统。
第二代逆向标记系统采用CVS-N2c毒株,它具有更低的神经毒性以及更强的跨突触突触传递能力,第二代逆向标记RV-CVS-N2C(G)-ΔG产品能在相同滴度达到比第一代RV-SAD-B19G-ΔG更好的逆向标记效果,且神经毒性更低。
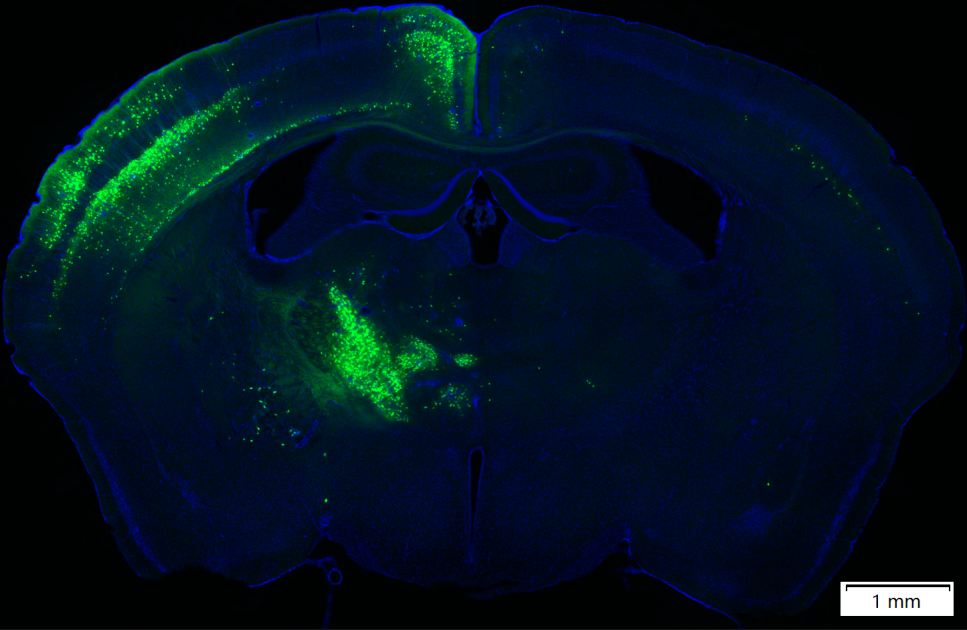
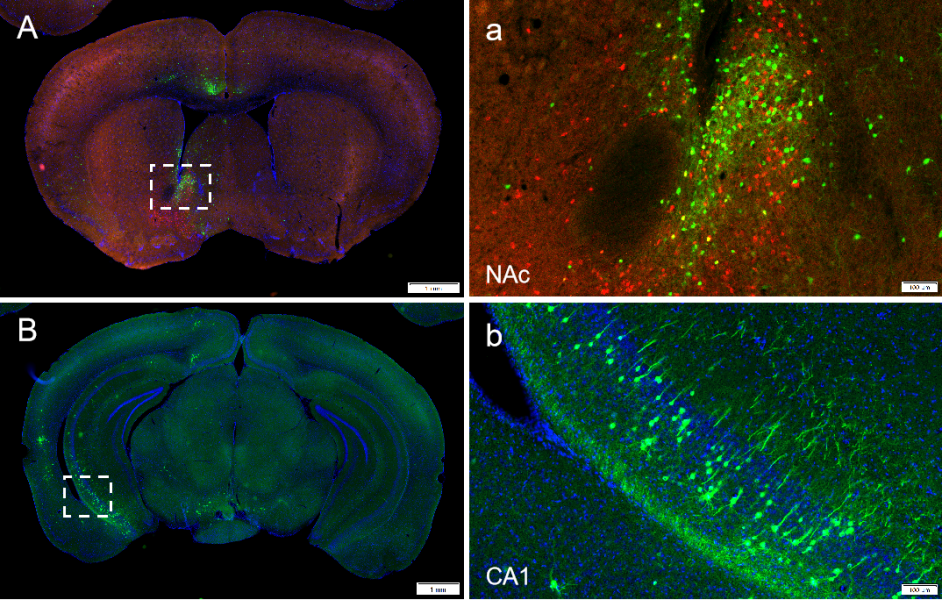
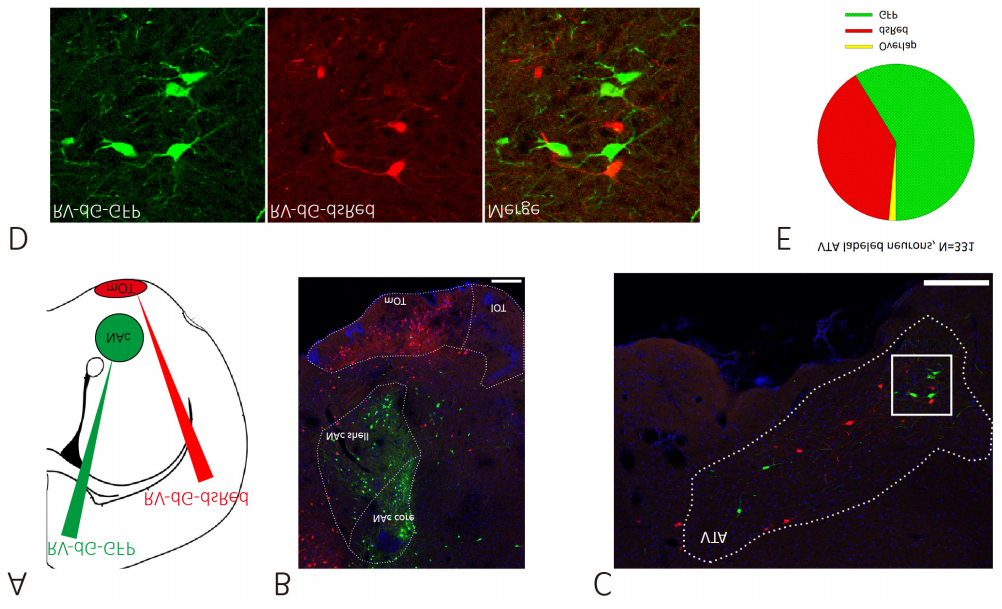
图3. RV-CVS-N2c-ΔG-EGFP(布林凯斯测试)
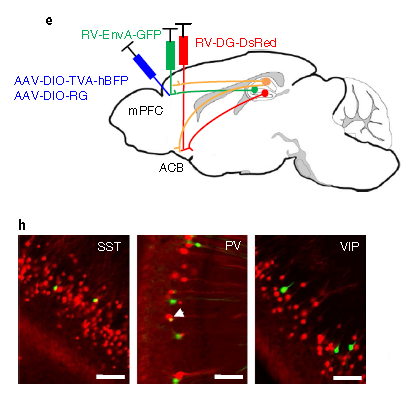
利用重组的禽类肉瘤病毒(Aviansarcoma-leukosisvirus,ASLV)的外膜蛋白(EnvA)融合蛋白包装RV-SAD-ΔG基因组形成的病毒粒子(RV-SAD-EnvA-ΔG),可特异性识别EnvA的受体TVA(TVA只存在于禽类细胞中,在哺乳动物神经元中无表达)介导病毒特异性地感染细胞。利用Cre转基因鼠结合Cre-LoxP控制表达TVA及G蛋白的AAV辅助病毒(DIO-TVA和DIO-RVG),可实现只在特定区域特异类型神经元中表达TVA及G蛋白,从而利用RV-SAD-EnvA-ΔG实现对特异类型神经元的逆向跨单级突触标记(如图3)。在此基础上,使RV-SAD-EnvA-ΔG带上ChR2及GCaMP等基因,可实现对特异类型神经元的直接输出 网络进行功能操纵及活动监测。
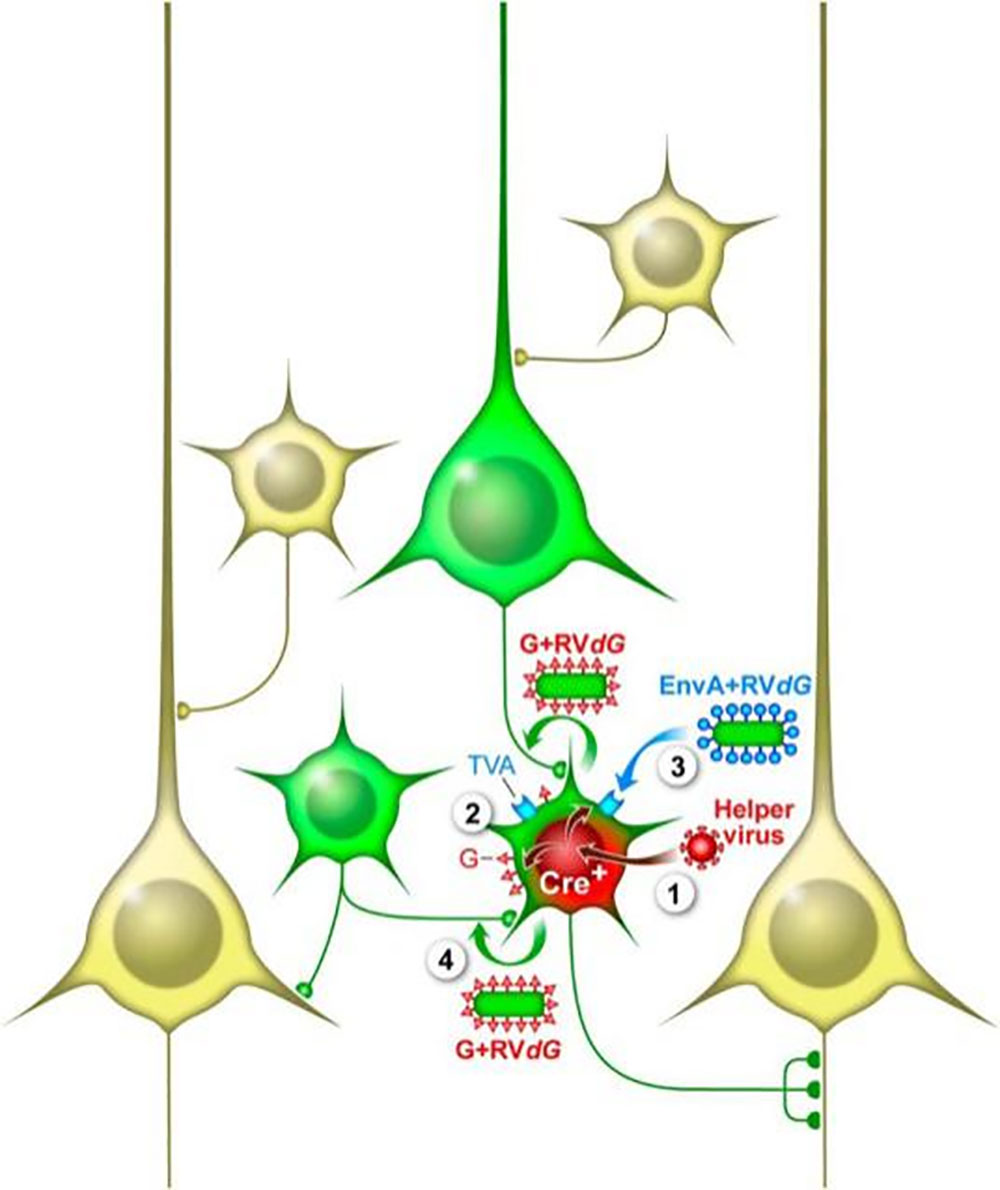
图4. RV-SAD-EnvA-ΔG实现神经元的逆向跨单级突触标记(Callaway, et al., The Journal of neuroscience: the official journal of the Society for Neuroscience, 2015)
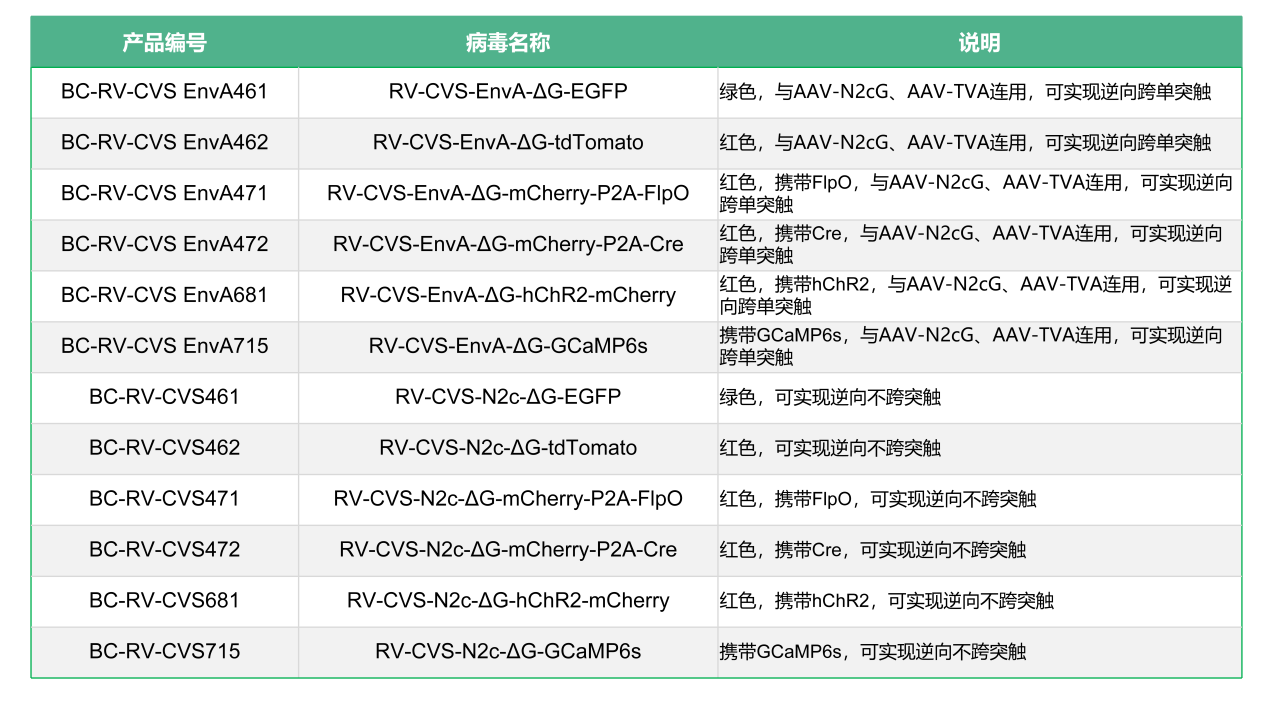
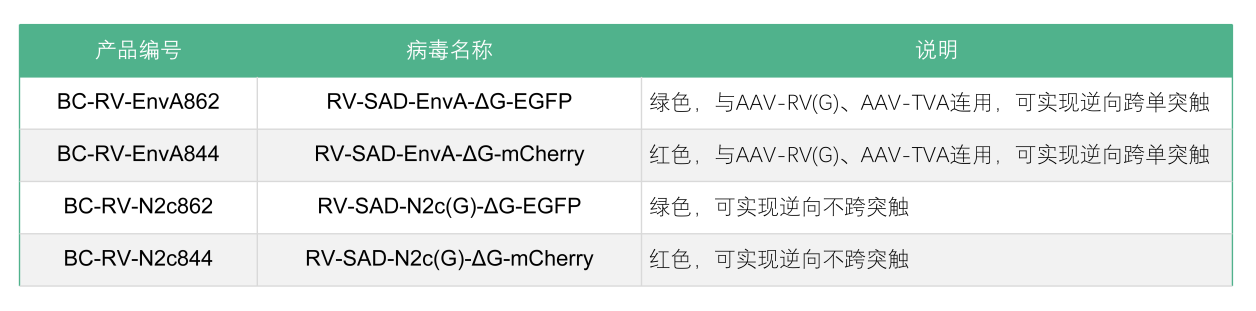
布林凯斯RV系列产品分为RV-CVS株和SAD-B19疫苗株。




地址:-
地址:-
