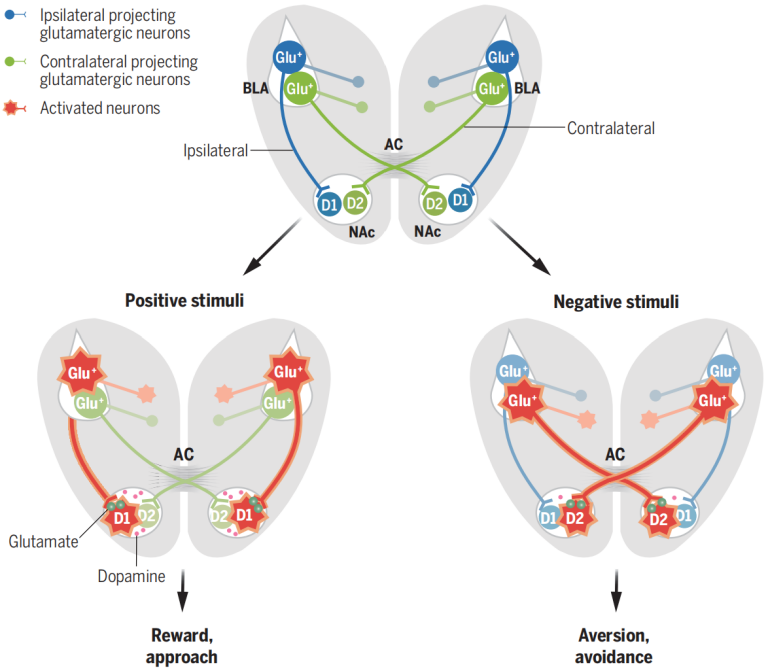
哺乳动物的大脑由两个半球组成,由三条主要的连合纤维连接到中线,分别是胼胝体(Corpus callosum,CC)、海马连合(Hippocampal commissure,HC)和前连合(Anterior commissure,AC),它们在生理上的重要性尚未被充分理解。前连合(AC)作为大脑中最古老的结构之一,在不同物种中的纤维组成差异较大,其在情绪和行为中的作用尚不清楚。基底外侧杏仁核(BLA)在编码正负情绪价值中扮演中心角色,且BLA通过AC向对侧半球的投射最近被发现,这提示了跨半球连通性在情绪处理中可能的重要性。BLA向伏隔核(NAc)发送谷氨酸能投射,而NAc是情绪和奖赏处理的关键脑区。研究者特别关注这些投射如何在正面和负面刺激之间产生不同的反应。

2024年11月8日,复旦大学刘星教授/马兰院士团队在Science杂志在线发表了题为“The interhemispheric amygdala-accumbens circuit encodes negative valence in mice”的研究论文,团队发现跨半球AC途径在处理负面信息和促进逃避行为中起着关键作用,这对于个体生存至关重要。对侧BLA-NAc投射与同侧投射在响应不同价值刺激、调节D1和D2-MSNs活动以及调节多巴胺释放方面具有不同的功能,其激活最终导致厌恶反应,揭示了跨半球BLA-AC-NAc环路在编码负面情绪中的关键作用。
在中线处切断了前连合(AC-cut;图1A),并检查了其行为效应。与未切断前连合(AC-uncut,假手术)和未经手术的(naïve)小鼠相比,AC-cut小鼠在运动活动、焦虑水平、社交能力或蔗糖偏好方面没有表现出差异。然而,前连合的切断似乎影响了厌恶反应。AC-cut小鼠比AC-uncut和naïve小鼠喝更多的奎宁溶液(引起社交回避)(图1,B和C)。在视觉悬崖测试(评估小鼠在面对潜在威胁时的行为选择,如偏好浅侧-安全区域-或深侧-危险区域-)中,与AC-uncut和naïve小鼠相比,AC-cut小鼠对浅侧的偏好减少(图1,D和E),冻结的时间很短,而大部分时间都在外部走动(图1,F和G)。此外,在社交回避测试中,AC-cut小鼠比AC-uncut和naïve小鼠花费更多时间探索攻击性的CD1小鼠(图1,H和I)。结合LiCl条件性场所回避(CPA)测试(社交回避)和恐惧条件测试结果,说明通过AC的跨半球连接对于负面刺激引起的厌恶反应至关重要。
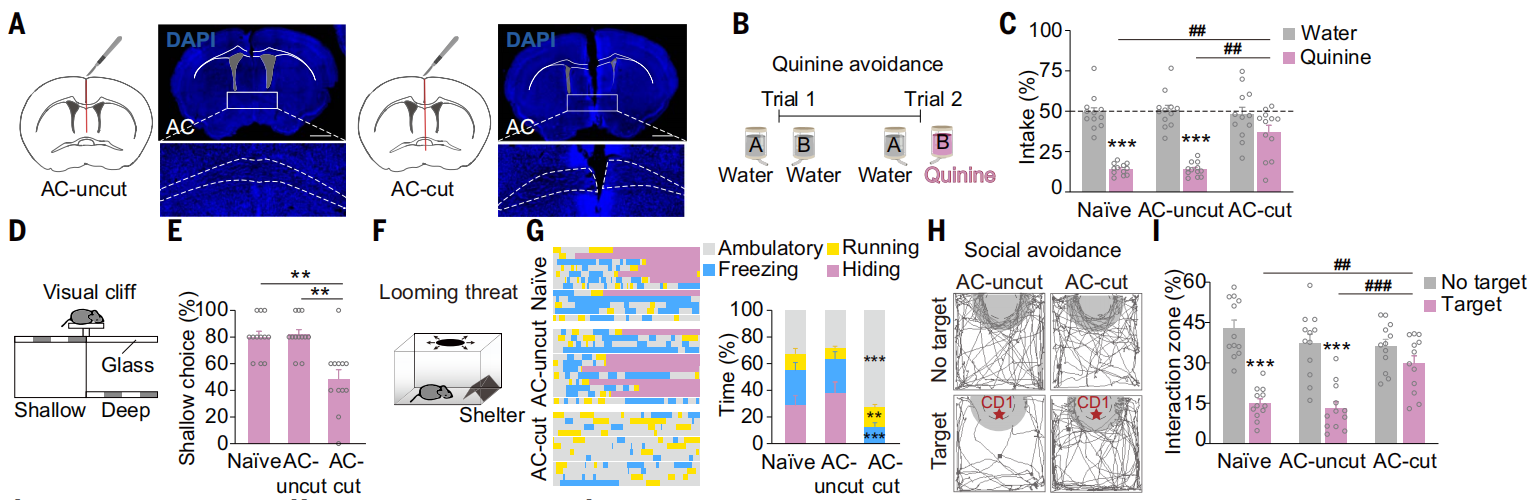
图1 AC切断导致厌恶情绪障碍
杏仁核在编码负面情绪值中扮演着关键角色,通过AC途径一些BLA神经元支配对侧的BLA。研究人员在单侧BLA谷氨酸能神经元中表达了mRuby,并使用X-CLARITY技术(一种先进的组织透明化和成像技术)追踪单侧BLA投射到整个大脑的路径(图2A)。三维(3D)重建图像揭示了三个主要的mRuby标记的投射起源于BLA(标记为I至III;图2A)。纹状终端(ST)途径(I)以倒U形从BLA延伸到同侧的NAc。腹侧杏仁核纤维(VAF)途径(II)与背侧ST途径在同侧NAc处汇聚,值得注意的是,另一腹侧轴突束(III)横向延伸至VAF途径,进入AC,并到达对侧半球(图2A和B)。
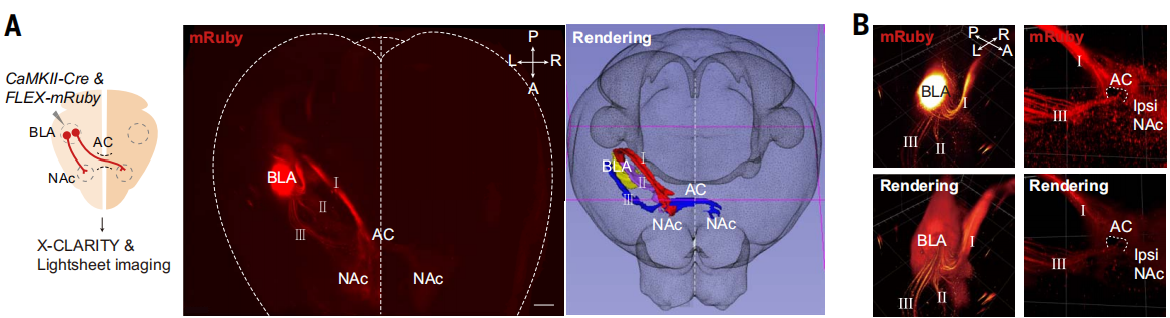
图2 BLA谷氨酸能神经元的全脑投射三维成像
单侧注射AAV到C57BL/6J小鼠BLA来介导ChR2-mCherry表达,在对侧和同侧NAc可以检测到BLA投射信号(图3C)。单侧注射携带Cre的顺行跨突触传递AAV到Ai14小鼠BLA,在同侧和对侧NAc中检测到tdTomato+细胞,其中细胞密度最高的在NAc核心区域(图3D)。随后,单侧注射CaMKII-Cre或GAD-Cre与floxed ChR2-mCherry的AAV混合液到小鼠BLA,观察到对侧NAc中有谷氨酸能投射,但没有观察到γ-氨基丁酸能投射(图3E和F)。此外,使用mGFP顺行标记显示,在中线处切断AC后,未检测到对侧BLA-NAc谷氨酸能投射。这些结果表明BLA的谷氨酸能神经元通过AC途径投射到对侧NAc。
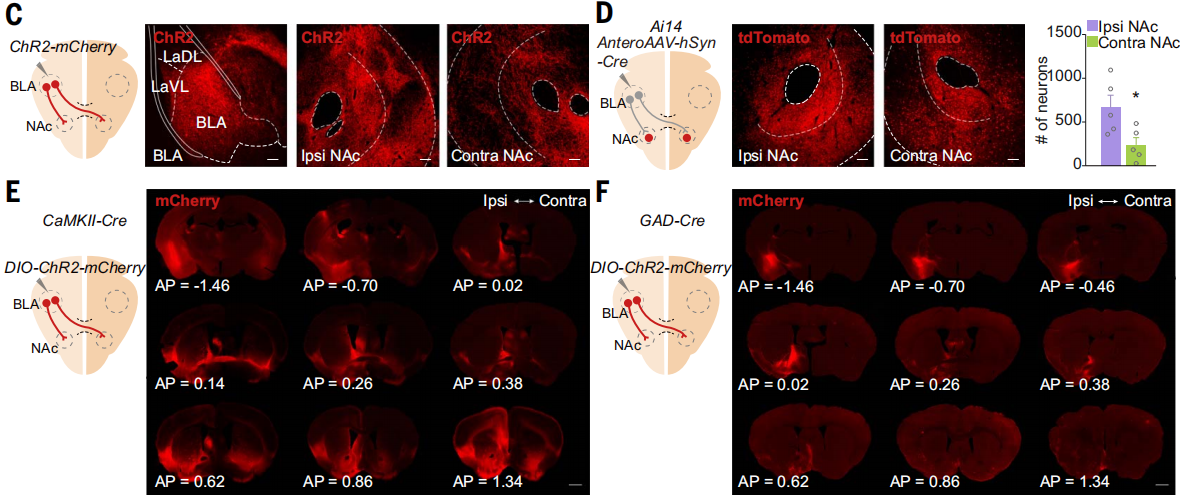
图3 BLA通过AC投射到对侧NAc
由于BLA谷氨酸能神经元可以投射到NAc,而NAc神经元不投射到BLA,研究人员在大脑一侧NAc注射逆行AAV-eGFP,另一侧NAc注射逆行AAV-tdTomato,以逆行标记投射到同侧和对侧NAc的BLA神经元,它们具有不同的荧光(图4I)。结果显示,67.50 ± 2.14%的BLA神经元投射到同侧NAc,36.98 ± 2.16%投射到对侧NAc,其中一小部分(4.48 ± 0.33%)双侧投射(图4J和K)。同侧和对侧BLA-NAc投射神经元在BLA的中间和后部区域最为密集(图4K)。RV逆向跨单突触示踪揭示了投射到同侧NAc的BLA神经元接收了来自终纹床核(BNST)和内侧前额叶皮质(mPFC)的更多输入,而投射到对侧NAc的BLA神经元则接收了来自外侧内嗅皮层(LEC)和梨状皮质(Pir)的更多输入(图4,L和M)。
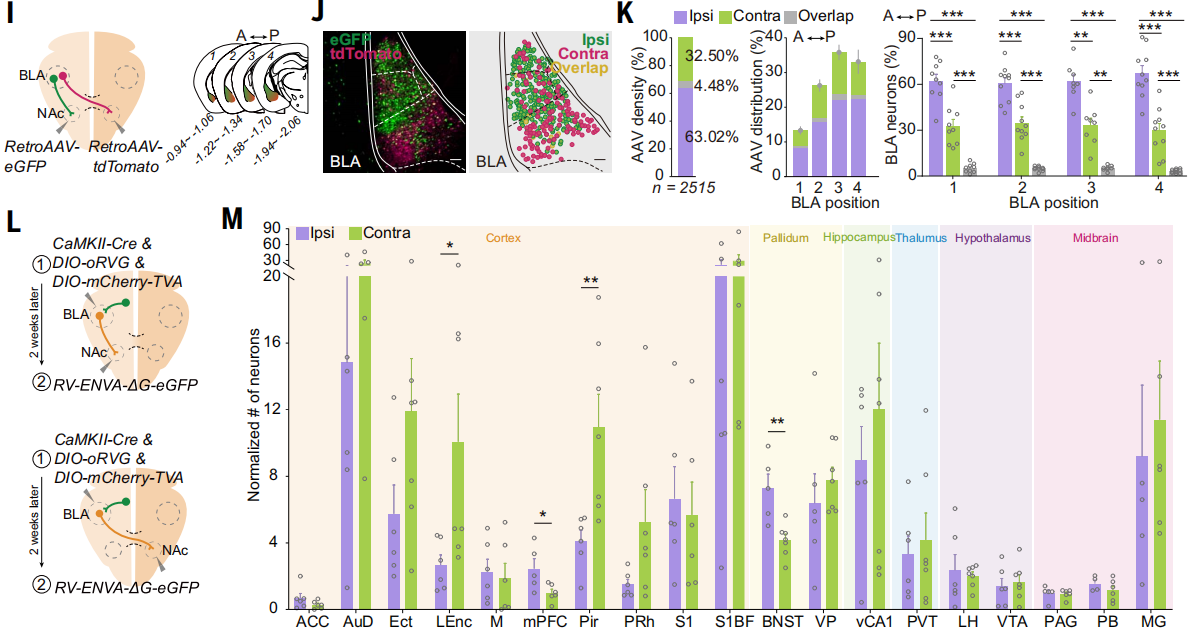
图4 同侧和对侧NAc投射的BLA神经元全脑输入区域量化
将AAV-Cre.on-FlpO.off-NBL10注射到一侧BLA,并将逆行AAV-Cre注射到同侧NAc,以及逆行AAV-FlpO注射到对侧NAc,或者反过来,将逆行AAV-FlpO注射到同侧NAc,逆行AAV-Cre注射到对侧NAc(图5A)。从投射同侧和对侧NAc的BLA神经元中分离出与核糖体相关的mRNA,并进行了测序(图5,A-C)。结果显示显示这两组神经元之间的基因表达存在显著差异:与同侧NAc投射的BLA神经元相比,对侧NAc投射的BLA神经元中有2107个基因的表达水平更高,2336个基因的表达水平更低(图5D)。京都基因与基因组百科全书(KEGG)对差异表达基因的分析结果显示多个通路相关的基因表达显著增加,对侧NAc投射的BLA神经元中的催产素信号途径,以及同侧NAc投射的BLA神经元中的轴突导向和谷氨酸能突触途径(图5E)。Glra3、Grm5、Ntsr2等主要在同侧NAc投射的BLA神经元中表达,而Oxtr、Adrb1等在对侧NAc投射的BLA神经元中富集(图5F)。
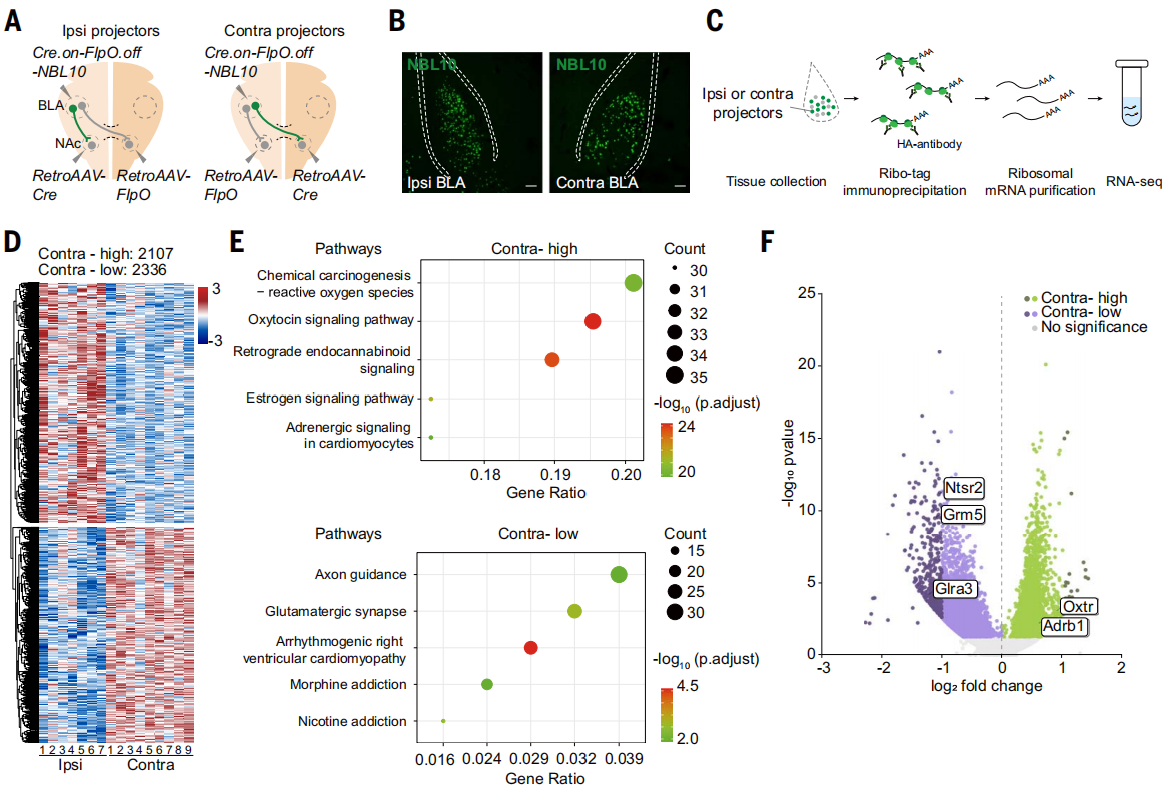
图5 同侧和对侧NAc投射的BLA神经元具有不同分子表型
为了检验对侧和同侧BLA-NAc投射对正面和负面刺激的反应,研究人员单侧注射AAV-CaMKIIa-Cre和AAV-DIO-axon-GCaMP6f到C57BL/6J小鼠的BLA,并记录了同侧和对侧NAc中BLA谷氨酸能投射纤维的钙信号变化(图6,A和B)。在自由消耗蔗糖溶液后,同侧BLA-NAc投射中检测到GCaMP荧光显著增加(图6,C-E)。相反,在舔舐奎宁溶液后,对侧BLA-NAc投射中发现了GCaMP荧光增加。这些结果表明,甜味和苦味分别激活了同侧和对侧BLA-NAc投射。嗅探一只陌生的雌性小鼠(社交奖励),增加了同侧BLA-NAc投射中GCaMP荧光。相反,逃离CD1攻击性小鼠的行为(逃跑行为),触发了对侧BLA-NAc投射中更大的钙离子信号变化(图6,F和G)。因此,社交奖励和社交回避分别激活了同侧和对侧的BLA-NAc投射。潜在威胁降低了同侧投射GCaMP荧光,但增加了对侧投射GCaMP荧光。此外,空气喷射和尾部悬挂引起的Ca2+瞬变在对侧投射中比同侧投射更高。对侧BLA-NAc投射被负面刺激激活并被正面刺激抑制。
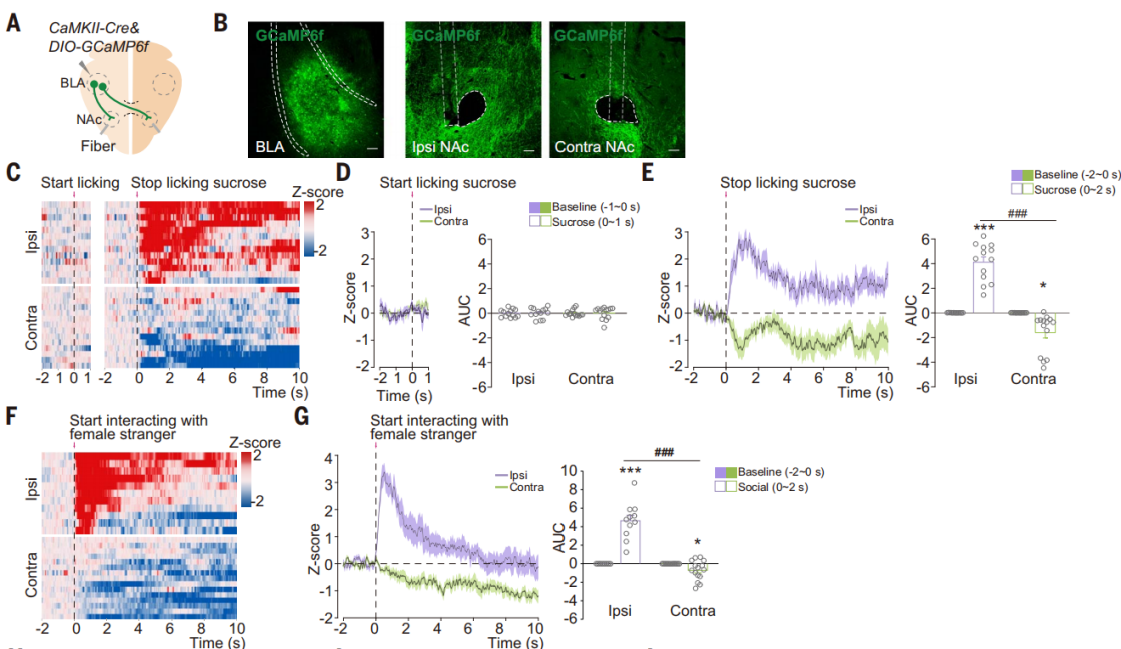
图6 同侧和对侧BLA-NAc投射被相反的刺激激活
研究人员将AAV-CaMKIIa-eNpHR3.0-mCherry注射到一侧BLA,将AAV-CaMKIIa-SwiChRca-EYFP注射到另一侧BLA,通过不同波长的激光选择性抑制投射到同侧或对侧NAc中轴突末梢(图7,A和B)。在蔗糖偏好测试或可卡因位置偏好测试中,选择性抑制同侧BLA-NAc投射减少了蔗糖摄入量(图7,C-E),并损害了可卡因条件性位置偏好(CPP)的获得(图7,H-J)。选择性抑制对侧BLA-NAc投射增加了奎宁消耗量(图7,F和G),在迫近威胁出现时减少了隐藏时间并增加了活动时间(图7,K和L),并减少了对CD1攻击性小鼠的回避。LiCl条件性场所回避(CPA)测试和恐惧条件测试中抑制对侧BLA-NAc投射也减少了对LiCl的位置厌恶或冻结行为。此外,在CPP装置中对小鼠进行了光遗传抑制同侧或对侧BLA-NAc投射的条件。数据显示,光遗传抑制同侧BLA-NAc投射条件的小鼠发展出了对激光配对侧的回避;相反,抑制对侧BLA-NAc投射的小鼠表现出对激光配对侧的偏好。以上结果说明,同侧和对侧的BLA-NAc投射通路分别参与小鼠正面情绪和负面情绪的行为。
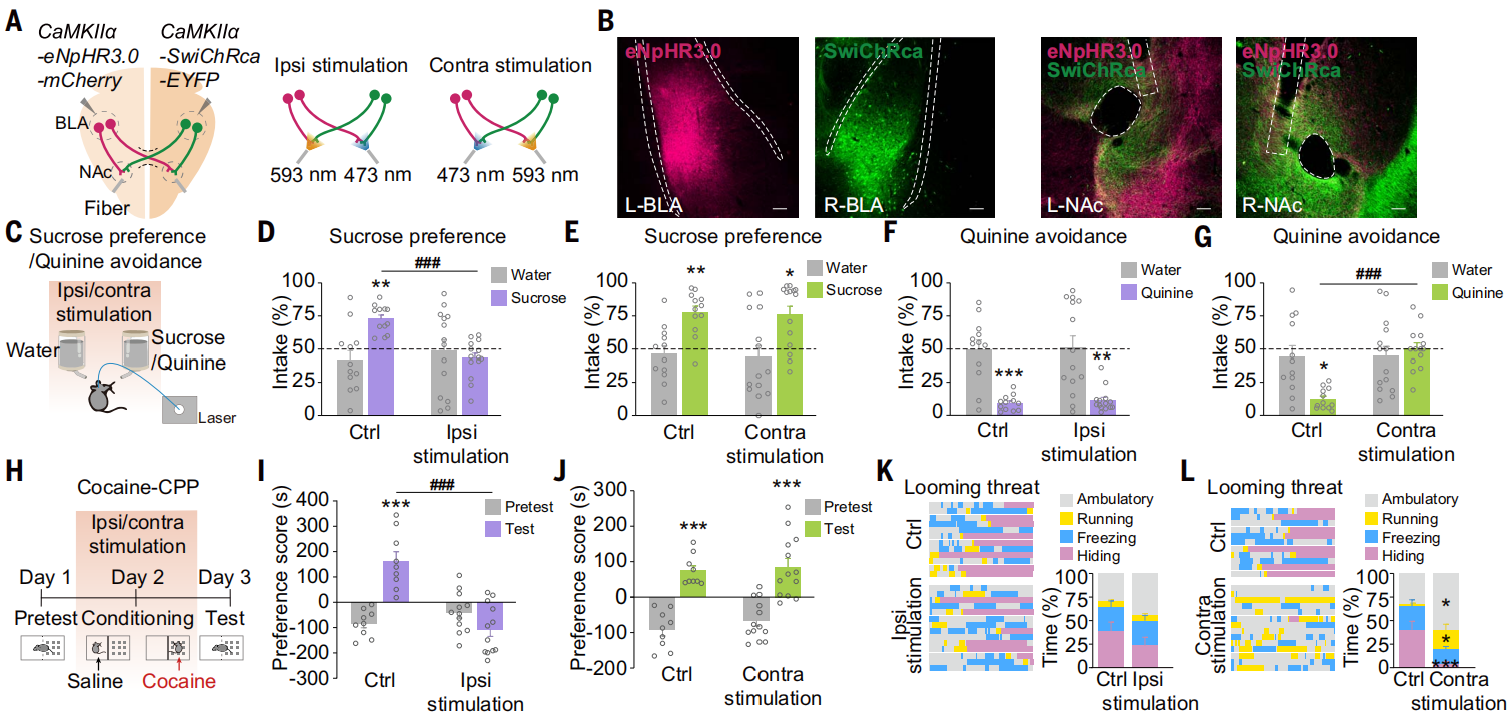
图7 对侧BLA-NAc投射编码负面情绪
刺激同侧和对侧BLA-NAc投射的是否会引发相反的情感价值状态?将AAV-CaMKIIa-ChrimsonR-mCherry注射到一侧BLA,将AAV-mCaMKIIa-hChR2-EYFP注射到另一侧BLA,以激活NAc中同侧或对侧的投射纤维(图8,A和B)。在实时位置偏好(RTPP)或回避(RTPA)测试中,激活同侧BLA-NAc投射增加小鼠停留在激光配对侧的时间,而激活对侧BLA-NAc投射效果相反(图8,C-H)。在RTPP-RTPA测试中,每只小鼠首先接受同侧BLA-NAc刺激,然后切换到对侧刺激。小鼠在同侧激活期间表现出强烈停留在激光配对侧的偏好,并在切换到对侧激活时转为回避激光配对侧,这表明激活对侧BLA-NAc可能抵消了同侧激活的效果(图8,I-K)。结合双孔激光颅内自我刺激(ICSS)实验(图8,L和M)和两侧BLA光遗传实验结果,进一步表明,激活同侧BLA-NAc投射介导正面情绪行为,而激活对侧BLA-NAc投射介导负面情绪行为,并且激活对侧的BLA-NAc能够拮抗激活同侧BLA-NAc的正面情绪行为。
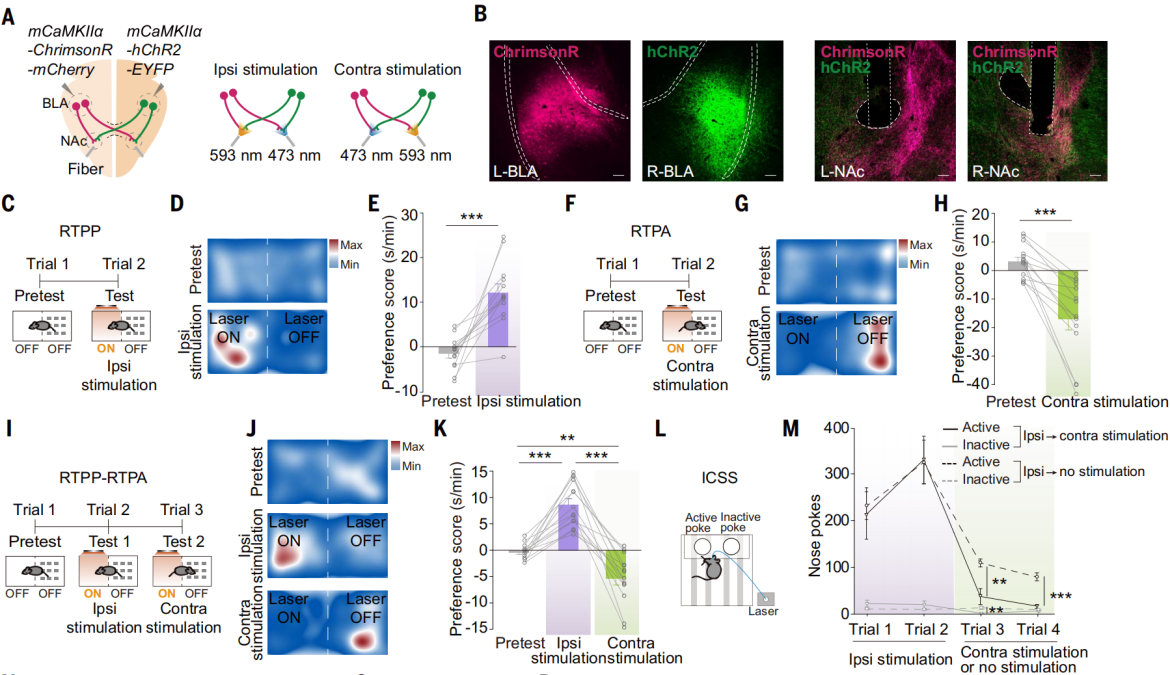
图8 激活同侧和对侧BLA-NAc投射诱导相反的行为模式
为了标记由同侧或对侧BLA支配的NAc神经元,研究人员向大脑一侧BLA注射顺行跨突触AAV1-FlpO,向另一侧BLA注射AAV1-Cre,同时向一侧NAc注射AAV-DIO-eGFP和AAV-fDIO-mCherry的混合物(图9A)。在标记的NAc神经元中,69.95 ± 4.41%只表达mCherry(仅来自同侧的输入),24.94 ± 4.01%只表达eGFP(仅来自对侧的输入),而只有5.11 ± 1.41%同时表达eGFP和mCherry(来自两侧的输入)(图9,B-D)。
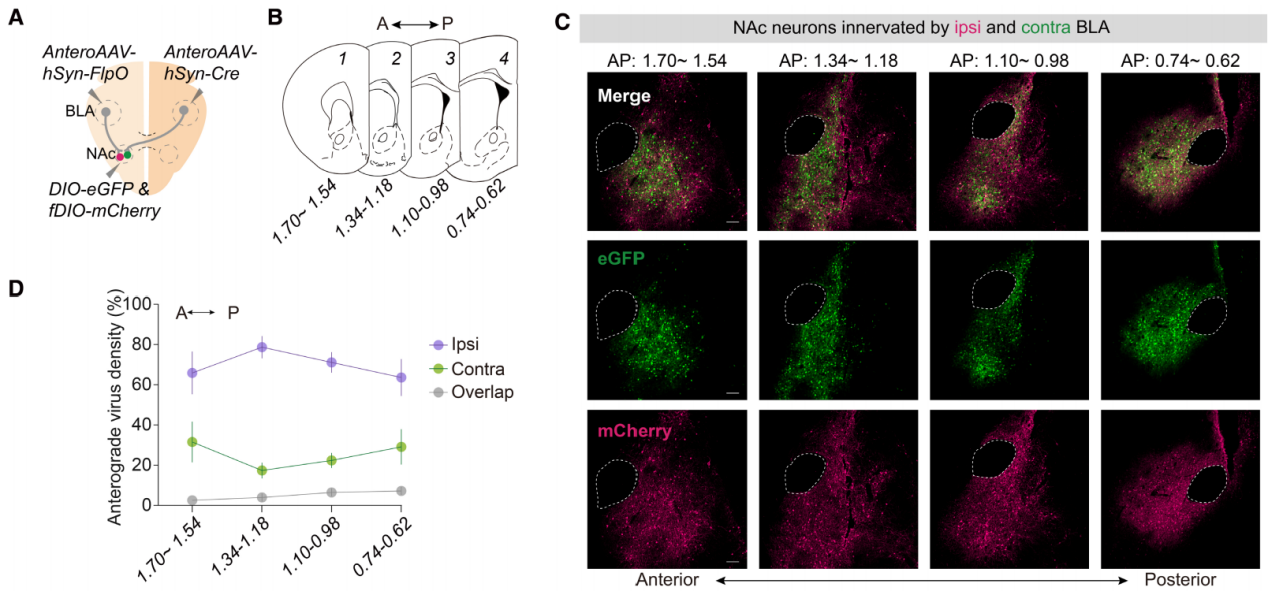
图9 标记同侧或对侧BLA支配的NAc神经元
多巴胺受体D1阳性和D2阳性的中棘神经元(D1-和D2-MSNs)对于情绪价值处理至关重要,它们构成了NAc神经元的大约95%。研究人员向一侧BLA注射顺行跨突触AAV1-Cre,并在D1-tdTomato小鼠双侧NAc注射AAV-DIO-eGFP(图10E)。结果显示,由同侧BLA-NAc投射支配的NAc神经元中有60.99 ± 2.00%为D1-MSNs,由对侧BLA-NAc投射支配的NAc神经元中有24.52 ± 1.61%为D1-MSNs(图10,F和G),在Ai14小鼠单侧BLA注射顺行跨突触AAV1-Cre,对NAc中被标记神经元进行smFISH分析进一步证实了这个结论(图10,H-J)。
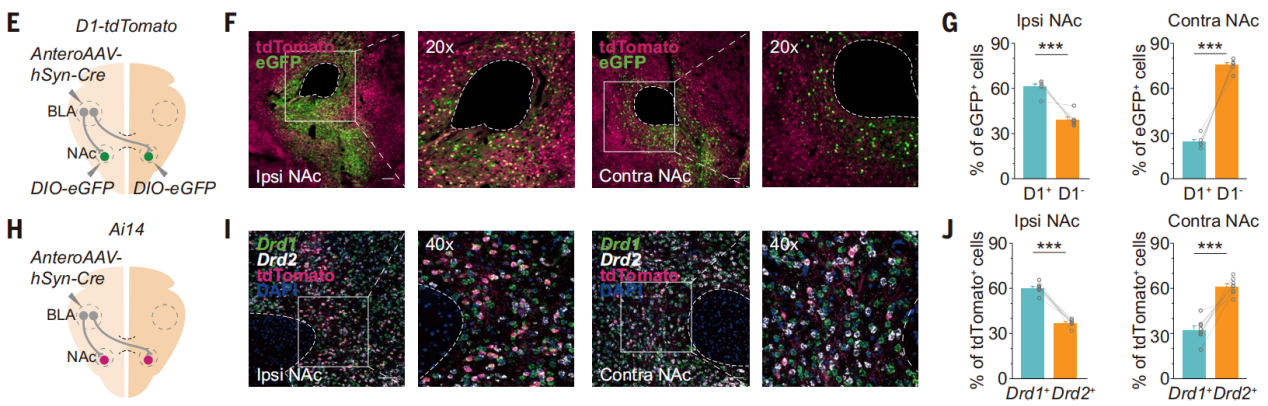
图10 同侧BLA-NAc投射支配NAc中的D1-MSNs神经元
PV、SOM和ChAT阳性神经元在NAc中占比不到10%,D2多巴胺受体也在NAc的ChAT内神经元中表达。光激活同侧BLA-NAc投射在D1-MSNs轴突末梢,发现GCaMP荧光的显著增加(图11,K-N),而光激活对侧BLA-NAc投射在D2-MSNs的轴突末梢,引起了GCaMP荧光的显著增加(图11,O-Q)。这些结果表明,同侧BLA-NAc投射主要支配NAc中的D1-MSNs,而对侧BLA-NAc投射主要支配NAc中的D2-MSNs。
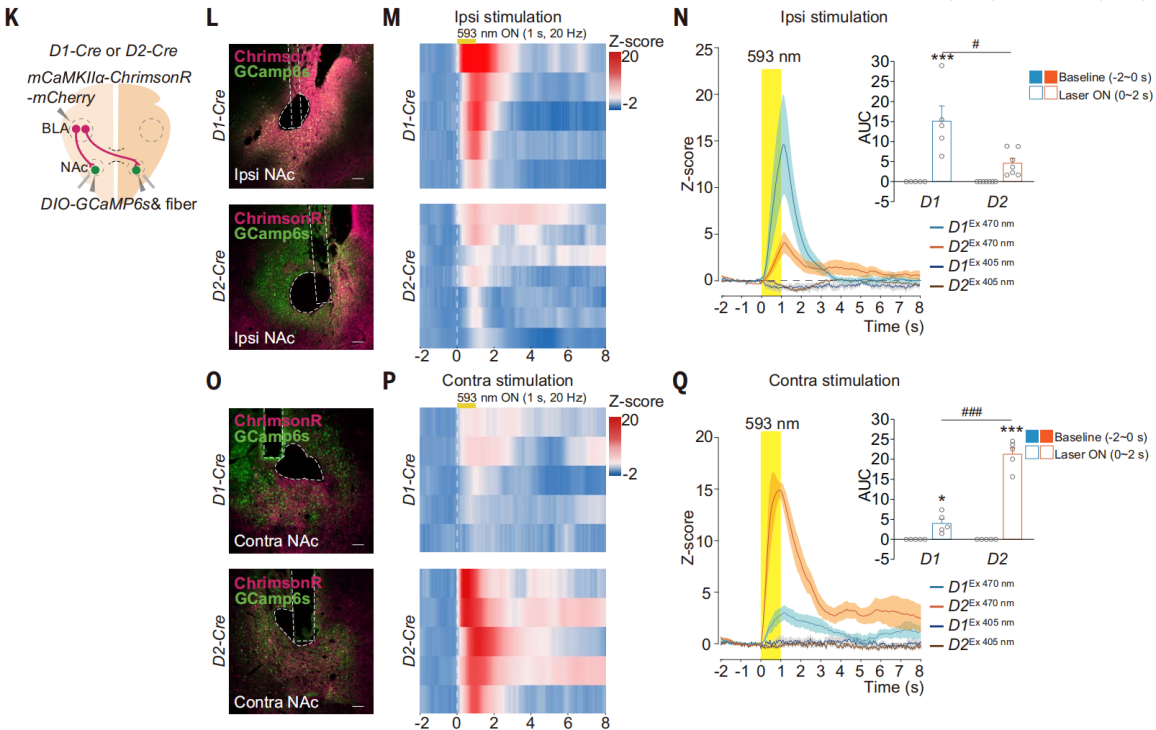
图11 对侧BLA-NAc投射支配NAc中的D2-MSNs神经元
为了进一步研究了BLA-NAc连接在调控情绪价值中作用,在D1-Cre或D2-Cre小鼠中注射AAV-mCaMKIIa-ChrimsonR-mCherry,并在一侧BLA中注射顺行跨突触scAAV-hSyn-FlpO,在同侧或对侧NAc注射AAV-EF1a-Cre.on-FlpO.on-hM4D-EYFP(图12,A和B)。数据显示,抑制由同侧BLA-NAc投射支配的D1-MSNs阻断了同侧BLA-NAc投射激活产生的RTPP,而抑制由对侧BLA-NAc投射支配的D2-MSNs阻止了对侧BLA-NAc投射激活诱导的RTPA(图12,C-G)。此外,抑制由同侧BLA-NAc投射支配的D1-MSNs损害了蔗糖偏好,而抑制由对侧BLA-NAc投射支配的D2-MSNs损害了奎宁回避(图12,H-N)。同侧BLA-NAc D1-MSN通路编码正面情绪,对侧BLA-NAc D2-MSN通路编码负面情绪。
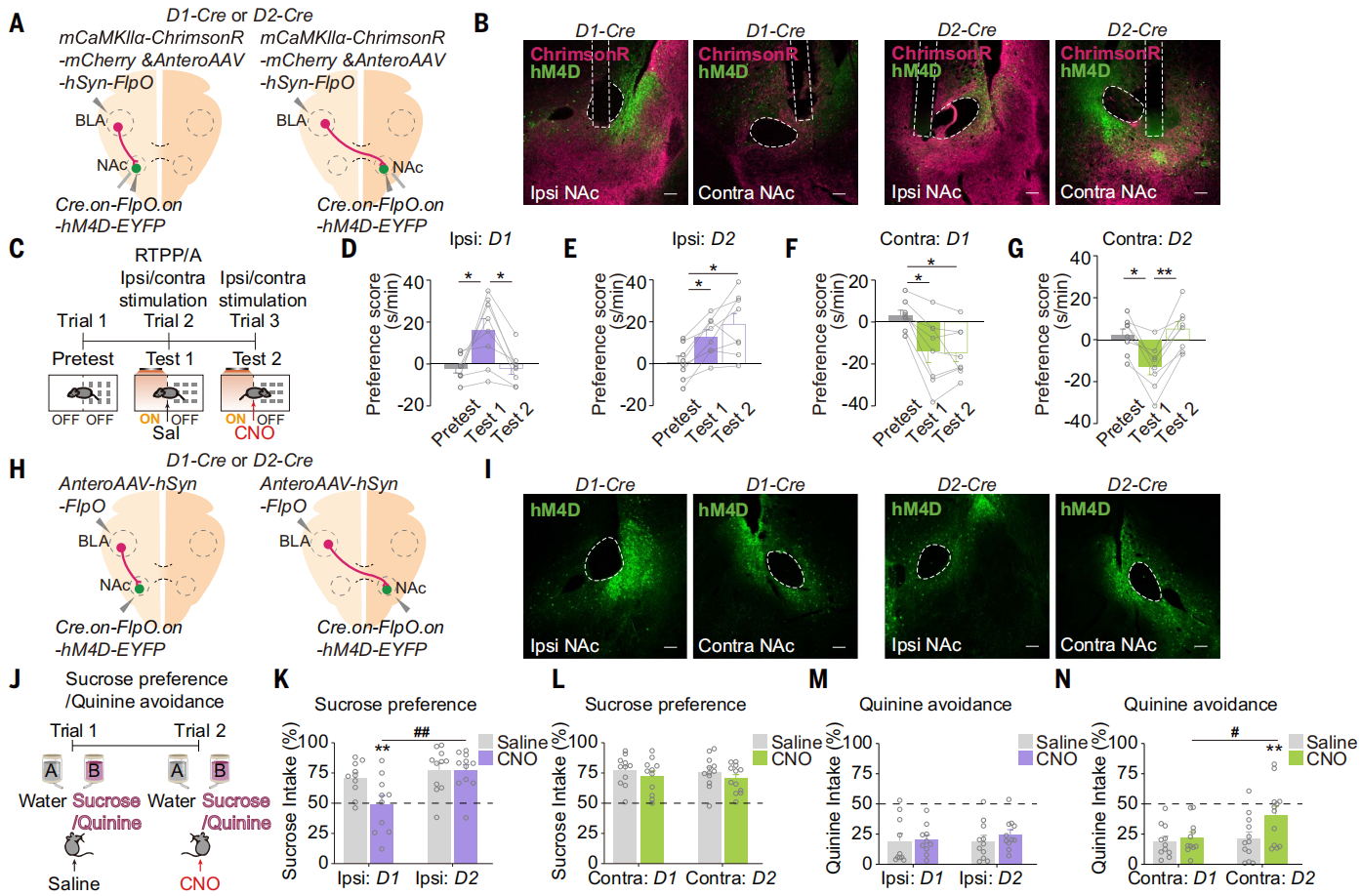
图12 同侧BLA-NAc D1-MSN通路编码正面情绪,对侧BLA-NAc D2-MSN通路路编码负面情绪
本文揭示了小鼠大脑中跨半球的BLA-NAc神经回路在编码负面情绪值中的关键作用,不仅增进了我们对情绪处理神经机制的理解,也为情绪相关疾病的诊断和治疗提供了新的科学依据,同时强调了跨半球信息传递在情绪和行为反应中的重要性。
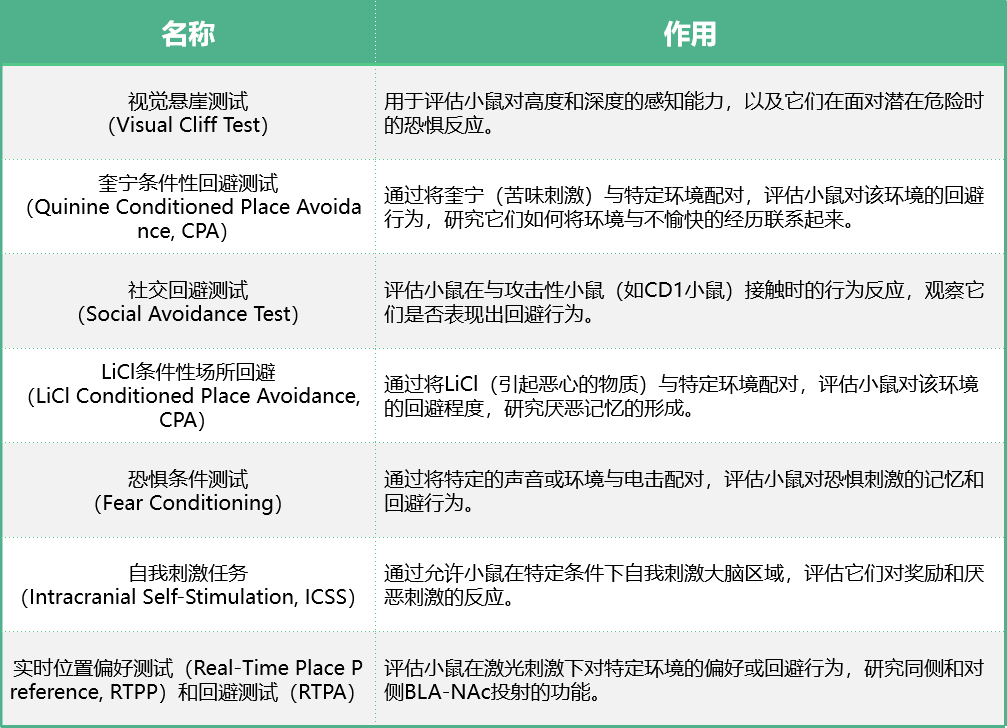
这篇文章中使用了多种行为学实验来研究小鼠对负面情绪和刺激的反应,具体包括以下几种实验:
本文未经授权禁止转载
如有需要,可联系小编获取原文




地址:-
地址:-
