IF: 26.8 | Nat. Biomed. Eng. | Pancreatic Neural Localization and Its Role in the Regulation of Glucose Metabolism
Release time:2024-12-31 16:52:36
The pancreas consists of both exocrine and endocrine components. The exocrine portion accounts for 95% of the pancreas's mass and produces digestive enzymes, while the endocrine portion is made up of islets, including β-cells and α-cells, which collectively maintain blood glucose levels. The pancreas is crucial for normal metabolism and digestion, with neural signaling playing a key role in regulating these functions. However, due to a lack of detailed understanding of neural inputs to the pancreas and the absence of tools specifically designed to modulate pancreatic neural functions, the precise role of pancreatic innervation in the regulation of metabolic and digestive processes remains insufficiently elucidated. Researchers utilized tissue clearing and retrograde tracing techniques, combined with targeted delivery of adeno-associated viruses (AAVs), to map the neural ganglionic inputs to the pancreas and validate the roles of parasympathetic and sympathetic pancreatic nerves in glucose regulation. They optimized the AAV serotypes, promoters, and delivery routes to specifically target pancreatic nerves. The study found that AAV8 and AAV9 preferentially transduce efferent neurons, while rAAV2-retro primarily targets afferent neurons. This research provides a method for precisely modulating specific pancreatic nerves to investigate their roles in metabolism and digestion, offering potential therapeutic strategies for pancreatic diseases such as diabetes, pancreatitis, and pancreatic cancer.
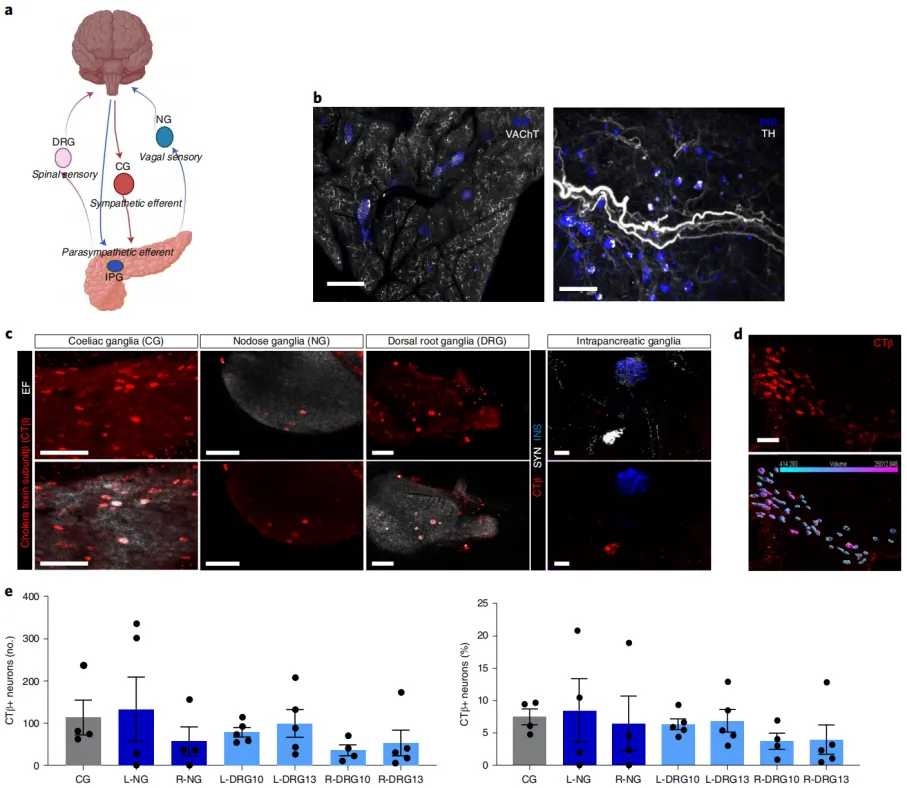
The pancreas is innervated by multiple peripheral ganglia.
Peripheral ganglia can regulate organ functions, integrate inputs, and potentially operate independently of central control. Therefore, characterizing the ganglia involved in pancreatic innervation provides crucial insights into its neural regulation. The pancreas is highly innervated by sympathetic ganglia (e.g., celiac ganglia, CG), parasympathetic ganglia (e.g., nodose ganglia, NG), and sensory nerves.
Using whole-organ tissue clearing techniques and specific neural markers, researchers confirmed a dense network of exocrine and endocrine neural fibers in the pancreas. Specifically, vesicular acetylcholine transporter (VAChT) was used to label efferent parasympathetic nerve fibers, while tyrosine hydroxylase (TH) was used to label efferent sympathetic nerve fibers.
To determine the projections of autonomic and sensory inputs to pancreatic innervation, non-viral tracers such as cholera toxin subunit B (CTB) were injected into the pancreas. Quantitative analysis was performed on CTB+ pancreatic projection neurons in extrinsic and intrinsic ganglia: CG, NG, and dorsal root ganglia (DRG) were categorized as extrinsic ganglia, while intrapancreatic ganglia were considered intrinsic.
The study revealed that in mice, the CG provided extensive projections to the pancreas. Sensory innervation primarily originated from sensory neurons of the vagus nerve and the spinal cord. Furthermore, the pancreas is innervated by distinct neuronal populations from the left and right sensory ganglia.
Figure 1: The pancreas is innervated by neurons from the celiac, nodose, dorsal root, and intrapancreatic ganglia.
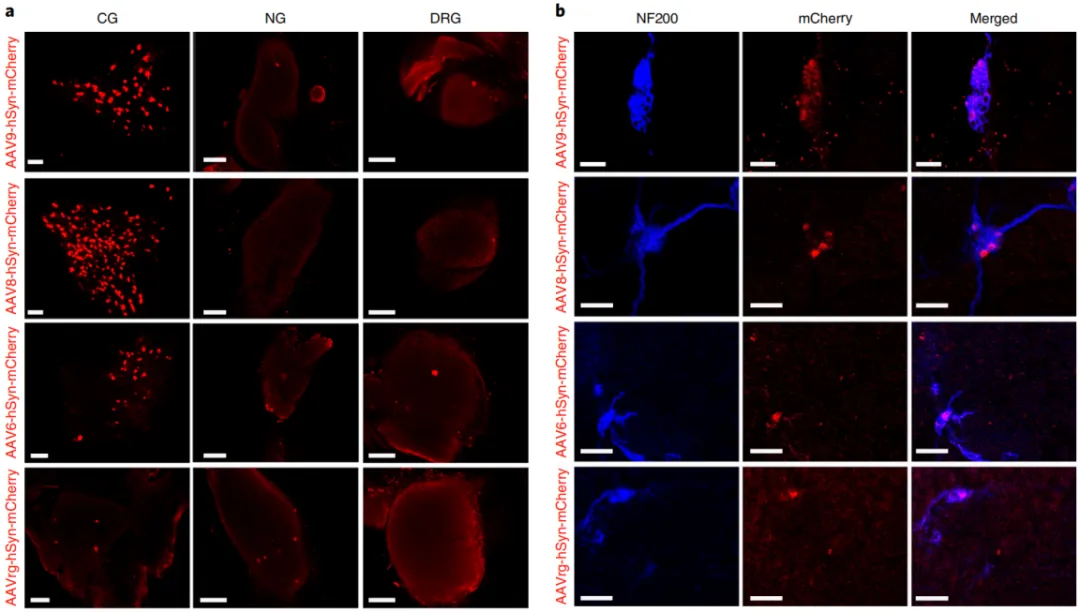
AAV selectively targets the pancreatic autonomic nerves.
Neurotracing using dyes and non-viral tracers such as CTB may experience some anterograde diffusion and excessive fluorescence intensity at the injection site, making it difficult to visualize short-distance neural connections, such as those in intrapancreatic ganglia. In contrast, viral tracers can provide specific retrograde tracing and bright long-distance connections by expressing fluorescent proteins. Therefore, AAV can be used to optimize retrograde viral tracing to better map the intrinsic and extrinsic ganglia that innervate the pancreas.
Researchers injected different AAV serotypes (including AAV6, AAV8, AAV9, and rAAV2-retro) into the pancreas to determine which serotypes effectively transduce pancreatic ganglia. The study found that AAV8 and AAV9 serotypes efficiently transduce efferent ganglia (such as CG and intrapancreatic ganglia), while rAAV2-retro primarily targets afferent ganglia (such as the NG), but with lower transduction efficiency. The AAV6 serotype showed low transduction efficiency in CG, NG, and DRG. Increasing the titer of AAV8 enhanced mCherry expression in the CG and intrapancreatic ganglia. By combining neuron-specific promoters and miR122 target sites, the researchers successfully restricted gene expression to pancreatic ganglia, reducing gene expression in non-target organs such as the liver.
Figure 2: AAV serotypes selectively target the pancreatic autonomic efferent and afferent nerves.
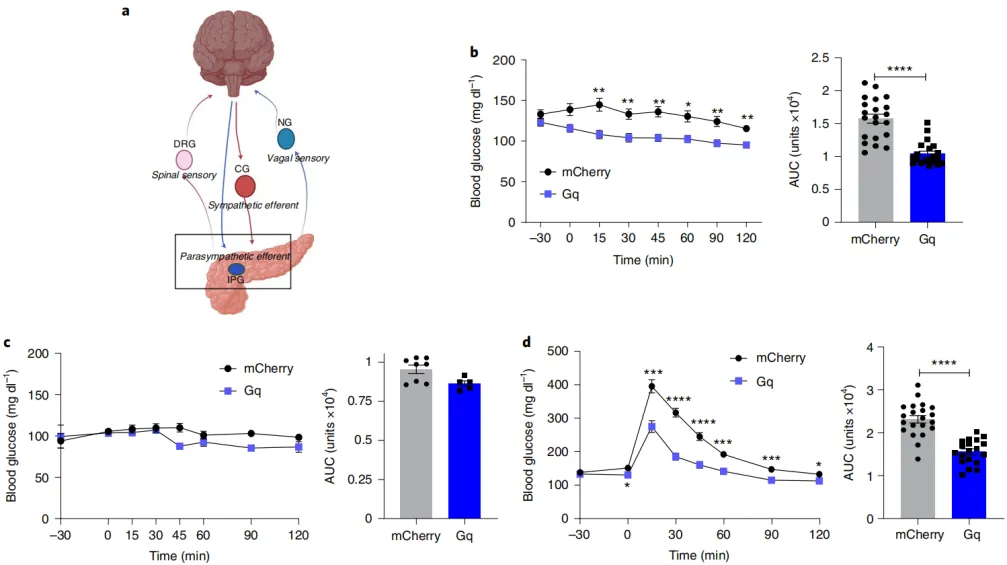
Activation of the pancreatic parasympathetic nerves can improve glucose regulation.
Next, the researchers applied the previously identified viral serotypes, promoters, and titers to achieve highly targeted regulation of pancreatic nerve innervation. The first step was to assess whether parasympathetic innervation of the pancreas was sufficient to regulate glucose homeostasis. In mice, the cell bodies of cholinergic efferent neurons in the pancreas are located in the intrapancreatic ganglia, and their fibers densely innervate pancreatic tissues, including the islets. Vagus nerve stimulation modulates both afferent and efferent fibers, while electrical stimulation and pharmacological methods simultaneously affect multiple metabolically active organs (liver, intestine, muscle, and fat).
To precisely investigate the physiological functions of parasympathetic efferent fibers in the pancreas, the researchers used optimized serotypes, promoters, viral titers, and delivery pathways, along with chemogenetic tools, to target specific neural networks. By using a dose of 5 times the usual amount of bile duct perfusion, they delivered a Cre-dependent activator hM3D(Gq) chemogenetic construct (AAV-Syn-DIO-hM3D(Gq)-mCherry) with AAV8 serotype to ChAT-IRES-CRE mice, thereby labeling cholinergic neurons in the intrapancreatic ganglia. The data indicated that selective activation of parasympathetic efferent innervation of the pancreas significantly increased plasma insulin and improved glucose tolerance in male mice without exacerbating hypoglycemia.
Figure 3: Activation of the pancreatic parasympathetic nerves can improve glucose regulation.
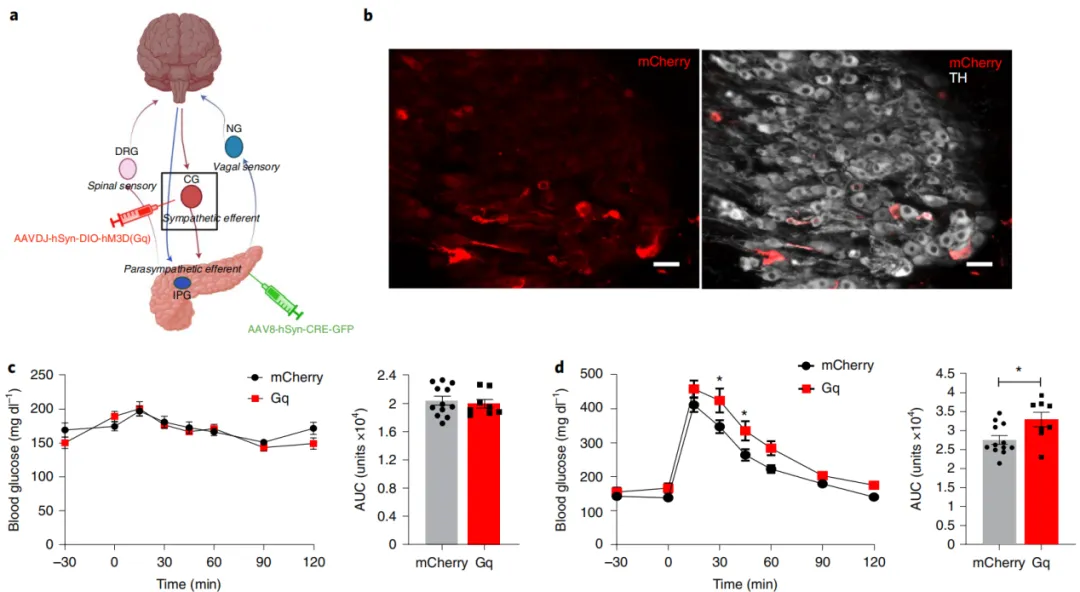
Activation of the pancreatic sympathetic nerves disrupts glucose homeostasis.
Previous studies have shown that stimulating the celiac nerve can increase glucagon and inhibit insulin release. However, this method regulates the pre-ganglionic sympathetic input to the celiac ganglia, affecting multiple organs within the celiac region. The celiac nerve also carries sensory afferent signals from various organs, making it impossible to isolate the effects of efferent sympathetic nerve activation on pancreatic function, even in in vitro studies.
To determine the role of sympathetic ganglia projecting to the pancreas in glucose control, researchers employed a dual-virus approach to specifically activate the sympathetic neurons projecting to the pancreas. They injected AAV8-hSyn-cre-GFP serotype into the pancreas to deliver Cre recombinase to the neurons in the CG that project to the pancreas, and simultaneously injected AAVdj-hSyn-DIO-hM3D(Gq)-mCherry or hSyn-DIO-mCherry into the celiac region to specifically express hM3D(Gq) or mCherry in the sympathetic neurons projecting to the pancreas.
The results showed that hM3D(Gq)-mCherry expression was found in sympathetic neurons in the CG, but not in the intrapancreatic ganglia. Activation of the sympathetic nerves projecting to the pancreas with CNO significantly impaired glucose tolerance. Compared to the control group injected with AAV8-hSyn-DIO-mCherry, the area under the curve (AUC) increased by 20%, with no impact on fasting blood glucose. In the insulin tolerance test, targeted activation of the sympathetic nerves projecting to the pancreas did not alter blood glucose levels.
Since one subset of sympathetic neurons projects to both the pancreas and the liver, the researchers used a pyruvate tolerance test to assess the impact of activating pancreatic-projecting sympathetic neurons on hepatic gluconeogenesis. In the pyruvate tolerance test, there was no difference in blood glucose between the hM3D(Gq)-mCherry expression group and the control group during the test.
Figure 4: Activation of the pancreatic sympathetic nerves disrupts glucose homeostasis.
Conclusion
By precisely mapping and specifically activating pancreatic ganglia, the roles of the parasympathetic and sympathetic nerves in regulating blood glucose homeostasis can be clearly defined. This not only enhances our understanding of the pancreatic neural regulation mechanisms but also provides new strategies for developing therapies targeting specific neural pathways, which has potential significant clinical value for treating metabolic diseases such as diabetes.